

GATA-2 reinforces megakaryocyte development in the absence of GATA-1
- PMID: 19620289
- PMCID: PMC2738300
- DOI: 10.1128/MCB.00482-09
GATA-2 reinforces megakaryocyte development in the absence of GATA-1
Abstract
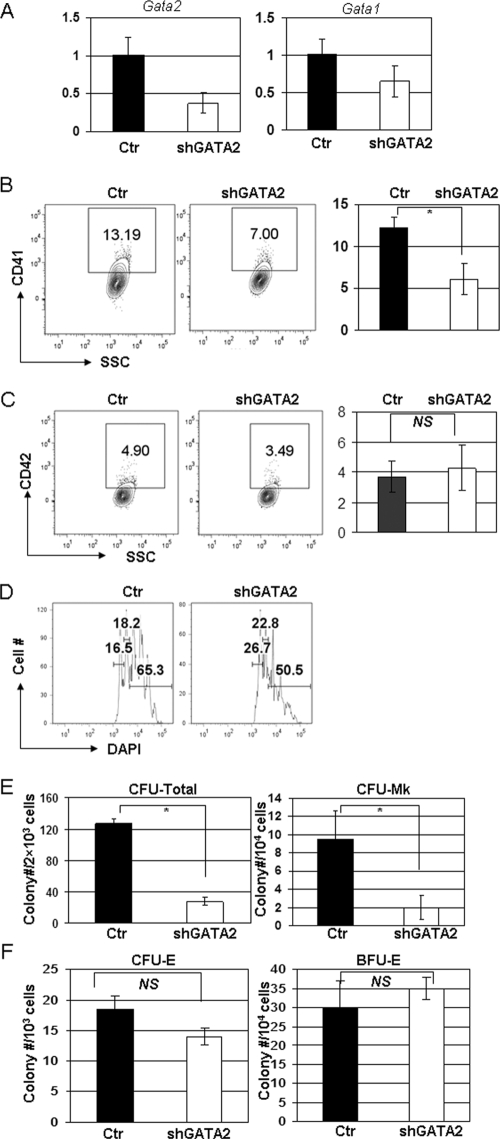
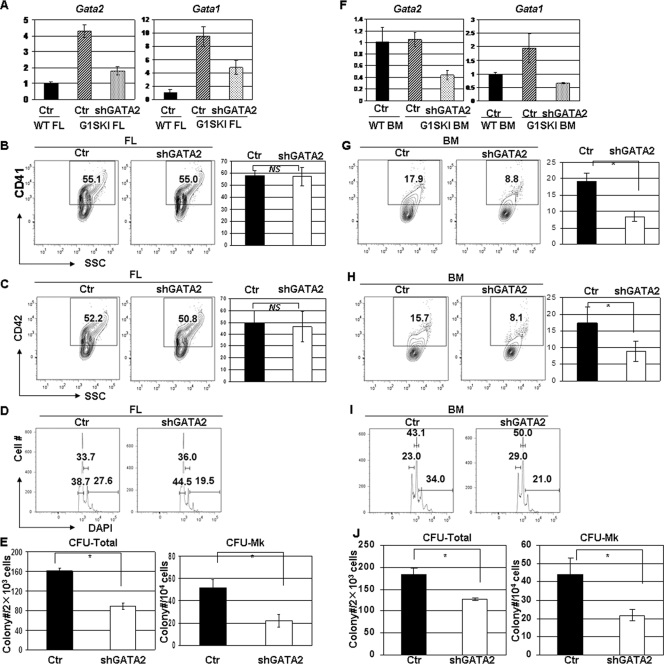
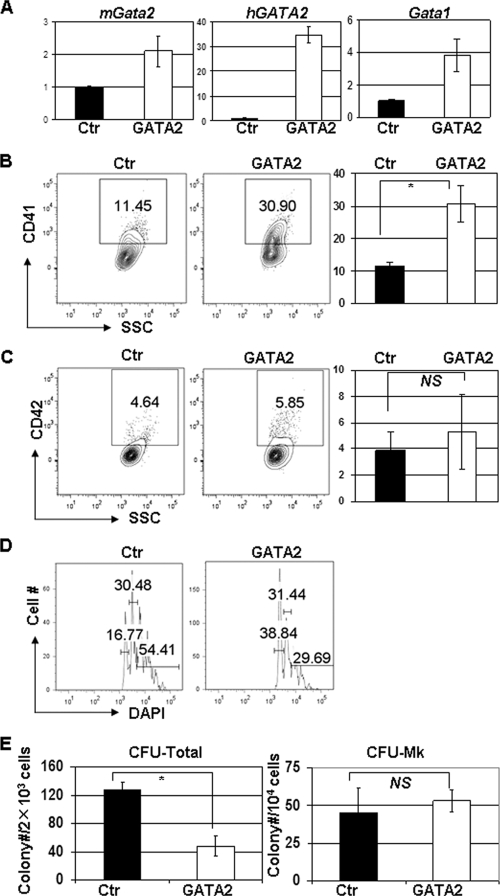
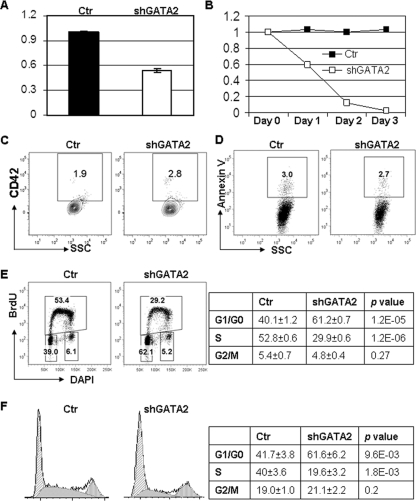
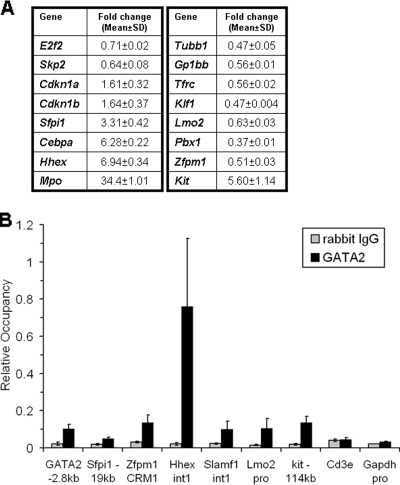
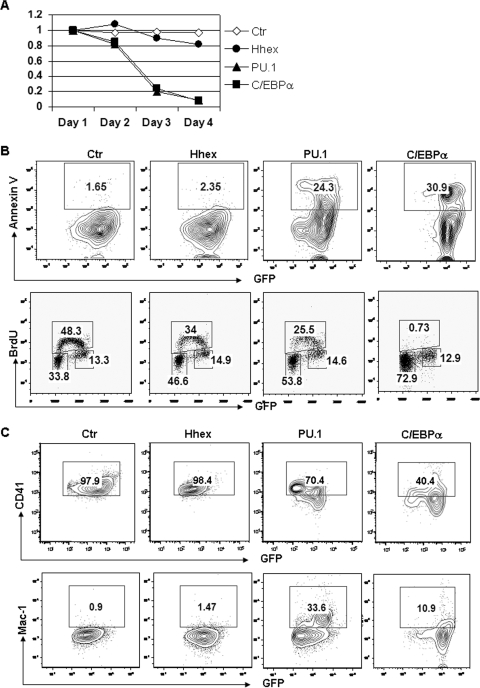
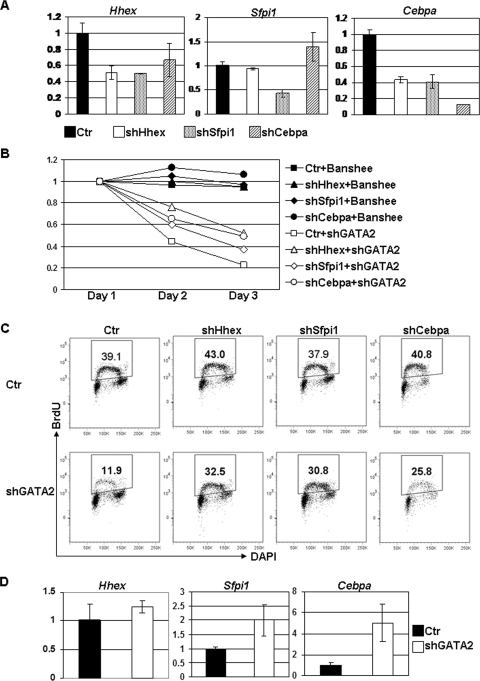
GATA-2 is an essential transcription factor that regulates multiple aspects of hematopoiesis. Dysregulation of GATA-2 is a hallmark of acute megakaryoblastic leukemia in children with Down syndrome, a malignancy that is defined by the combination of trisomy 21 and a GATA1 mutation. Here, we show that GATA-2 is required for normal megakaryocyte development as well as aberrant megakaryopoiesis in Gata1 mutant cells. Furthermore, we demonstrate that GATA-2 indirectly controls cell cycle progression in GATA-1-deficient megakaryocytes. Genome-wide microarray analysis and chromatin immunoprecipitation studies revealed that GATA-2 regulates a wide set of genes, including cell cycle regulators and megakaryocyte-specific genes. Surprisingly, GATA-2 also negatively regulates the expression of crucial myeloid transcription factors, such as Sfpi1 and Cebpa. In the absence of GATA-1, GATA-2 prevents induction of a latent myeloid gene expression program. Thus, GATA-2 contributes to cell cycle progression and the maintenance of megakaryocyte identity of GATA-1-deficient cells, including GATA-1s-expressing fetal megakaryocyte progenitors. Moreover, our data reveal that overexpression of GATA-2 facilitates aberrant megakaryopoiesis.
Figures

Similar articles
-
PSTPIP2 dysregulation contributes to aberrant terminal differentiation in GATA-1-deficient megakaryocytes by activating LYN.Cell Death Dis. 2014 Jan 9;5(1):e988. doi: 10.1038/cddis.2013.512. Cell Death Dis. 2014. PMID: 24407241 Free PMC article.
-
FIP1L1-PDGFRalpha imposes eosinophil lineage commitment on hematopoietic stem/progenitor cells.J Biol Chem. 2009 Mar 20;284(12):7719-32. doi: 10.1074/jbc.M807489200. Epub 2009 Jan 14. J Biol Chem. 2009. PMID: 19147501 Free PMC article.
-
Graded repression of PU.1/Sfpi1 gene transcription by GATA factors regulates hematopoietic cell fate.Blood. 2009 Jul 30;114(5):983-94. doi: 10.1182/blood-2009-03-207944. Epub 2009 Jun 2. Blood. 2009. PMID: 19491391 Free PMC article.
-
GATA switches as developmental drivers.J Biol Chem. 2010 Oct 8;285(41):31087-93. doi: 10.1074/jbc.R110.159079. Epub 2010 Jul 29. J Biol Chem. 2010. PMID: 20670937 Free PMC article. Review.
-
GATA-related hematologic disorders.Exp Hematol. 2016 Aug;44(8):696-705. doi: 10.1016/j.exphem.2016.05.010. Epub 2016 May 25. Exp Hematol. 2016. PMID: 27235756 Review.
Cited by
-
The Hematopoietic Stem and Progenitor Cell Cistrome: GATA Factor-Dependent cis-Regulatory Mechanisms.Curr Top Dev Biol. 2016;118:45-76. doi: 10.1016/bs.ctdb.2016.01.002. Epub 2016 Feb 26. Curr Top Dev Biol. 2016. PMID: 27137654 Free PMC article. Review.
-
Distinct, strict requirements for Gfi-1b in adult bone marrow red cell and platelet generation.J Exp Med. 2014 May 5;211(5):909-27. doi: 10.1084/jem.20131065. Epub 2014 Apr 7. J Exp Med. 2014. PMID: 24711581 Free PMC article.
-
Role of MicroRNAs and their Downstream Target Transcription Factors in Zebrafish Thrombopoiesis.Res Sq [Preprint]. 2023 Apr 24:rs.3.rs-2807790. doi: 10.21203/rs.3.rs-2807790/v1. Res Sq. 2023. Update in: Sci Rep. 2023 Sep 26;13(1):16066. doi: 10.1038/s41598-023-42868-7 PMID: 37162944 Free PMC article. Updated. Preprint.
-
Beta-Actin Is Involved in Modulating Erythropoiesis during Development by Fine-Tuning Gata2 Expression Levels.PLoS One. 2013 Jun 26;8(6):e67855. doi: 10.1371/journal.pone.0067855. Print 2013. PLoS One. 2013. PMID: 23840778 Free PMC article.
-
Effect of Mesenchymal Stem Cell-derived Microvesicles on Megakaryocytic Differentiation of CD34+ Hematopoietic Stem Cells.Adv Pharm Bull. 2020 Jun;10(2):315-322. doi: 10.34172/apb.2020.038. Epub 2020 Feb 20. Adv Pharm Bull. 2020. PMID: 32373502 Free PMC article.
References
-
- Bourquin, J. P., A. Subramanian, C. Langebrake, D. Reinhardt, O. Bernard, P. Ballerini, A. Baruchel, H. Cave, N. Dastugue, H. Hasle, G. L. Kaspers, M. Lessard, L. Michaux, P. Vyas, E. van Wering, C. M. Zwaan, T. R. Golub, and S. H. Orkin. 2006. Identification of distinct molecular phenotypes in acute megakaryoblastic leukemia by gene expression profiling. Proc. Natl. Acad. Sci. USA 1033339-3344. - PMC - PubMed
-
- Briegel, K., K. C. Lim, C. Plank, H. Beug, J. D. Engel, and M. Zenke. 1993. Ectopic expression of a conditional GATA-2/estrogen receptor chimera arrests erythroid differentiation in a hormone-dependent manner. Genes Dev. 71097-1109. - PubMed
-
- Cantor, A. B. 2005. GATA transcription factors in hematologic disease. Int. J. Hematol. 81378-384. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases