

A novel protein phosphatase 1-dependent spindle checkpoint silencing mechanism
- PMID: 19592249
- PMCID: PMC2791888
- DOI: 10.1016/j.cub.2009.05.060
A novel protein phosphatase 1-dependent spindle checkpoint silencing mechanism
Abstract
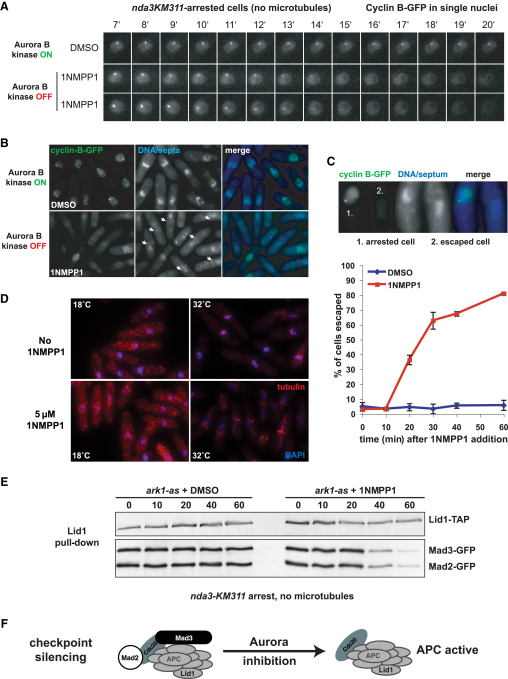
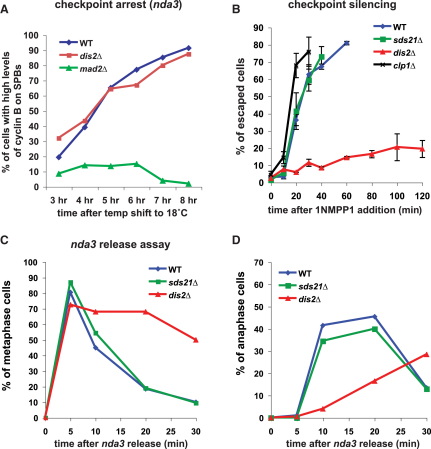
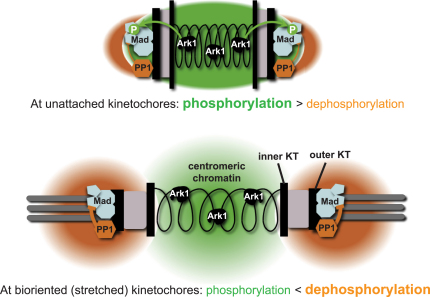
The spindle checkpoint is a surveillance system acting in mitosis to delay anaphase onset until all chromosomes are properly attached to the mitotic spindle. When the checkpoint is activated, the Mad2 and Mad3 proteins directly bind and inhibit Cdc20, which is an essential activator of an E3 ubiquitin ligase known as the anaphase-promoting complex (APC). When the checkpoint is satisfied, Cdc20-APC is activated and polyubiquitinates securin and cyclin, leading to the dissolution of sister chromatid cohesion and mitotic progression. Several protein kinases play critical roles in spindle checkpoint signaling, but the mechanism (or mechanisms) by which they inhibit mitotic progression remains unclear. Furthermore, it is not known whether their activity needs to be reversed by protein phosphatases before anaphase onset can occur. Here we employ fission yeast to show that Aurora (Ark1) kinase activity is directly required to maintain spindle checkpoint arrest, even in the presence of many unattached kinetochores. Upon Ark1 inhibition, checkpoint complexes are disassembled and cyclin B is rapidly degraded. Importantly, checkpoint silencing and cyclin B degradation require the kinetochore-localized isoform of protein phosphatase 1 (PP1(Dis2)). We propose that PP1(Dis2)-mediated dephosphorylation of checkpoint components forms a novel spindle checkpoint silencing mechanism.
Figures
Comment in
-
Cell division: righting the check.Curr Biol. 2009 Jul 28;19(14):R550-3. doi: 10.1016/j.cub.2009.06.047. Curr Biol. 2009. PMID: 19640489 Free PMC article. Review.
Similar articles
-
The spindle checkpoint functions of Mad3 and Mad2 depend on a Mad3 KEN box-mediated interaction with Cdc20-anaphase-promoting complex (APC/C).J Biol Chem. 2008 Aug 22;283(34):23039-47. doi: 10.1074/jbc.M803594200. Epub 2008 Jun 13. J Biol Chem. 2008. PMID: 18556659 Free PMC article.
-
Bub1 and aurora B cooperate to maintain BubR1-mediated inhibition of APC/CCdc20.J Cell Sci. 2005 Aug 15;118(Pt 16):3639-52. doi: 10.1242/jcs.02487. Epub 2005 Jul 26. J Cell Sci. 2005. PMID: 16046481
-
Kinase activity of fission yeast Mph1 is required for Mad2 and Mad3 to stably bind the anaphase promoting complex.Curr Biol. 2012 Feb 21;22(4):296-301. doi: 10.1016/j.cub.2011.12.049. Epub 2012 Jan 25. Curr Biol. 2012. PMID: 22281223 Free PMC article.
-
The spindle checkpoint: how do cells delay anaphase onset?SEB Exp Biol Ser. 2008;59:243-56. SEB Exp Biol Ser. 2008. PMID: 18368927 Review.
-
Cyclin A and Nek2A: APC/C-Cdc20 substrates invisible to the mitotic spindle checkpoint.Biochem Soc Trans. 2010 Feb;38(Pt 1):72-7. doi: 10.1042/BST0380072. Biochem Soc Trans. 2010. PMID: 20074038 Review.
Cited by
-
Microtubule attachment and spindle assembly checkpoint signalling at the kinetochore.Nat Rev Mol Cell Biol. 2013 Jan;14(1):25-37. doi: 10.1038/nrm3494. Nat Rev Mol Cell Biol. 2013. PMID: 23258294 Free PMC article. Review.
-
Recent Progress on the Localization of PLK1 to the Kinetochore and Its Role in Mitosis.Int J Mol Sci. 2022 May 8;23(9):5252. doi: 10.3390/ijms23095252. Int J Mol Sci. 2022. PMID: 35563642 Free PMC article. Review.
-
Regulated inactivation of the spindle assembly checkpoint without functional mitotic spindles.EMBO J. 2011 Jun 3;30(13):2648-61. doi: 10.1038/emboj.2011.176. EMBO J. 2011. PMID: 21642954 Free PMC article.
-
Spindle checkpoint silencing requires association of PP1 to both Spc7 and kinesin-8 motors.Dev Cell. 2011 Jun 14;20(6):739-50. doi: 10.1016/j.devcel.2011.05.008. Dev Cell. 2011. PMID: 21664573 Free PMC article.
-
Bub1-mediated adaptation of the spindle checkpoint.PLoS Genet. 2011 Jan 27;7(1):e1001282. doi: 10.1371/journal.pgen.1001282. PLoS Genet. 2011. PMID: 21298086 Free PMC article.
References
-
- Musacchio A., Salmon E.D. The spindle-assembly checkpoint in space and time. Nat. Rev. Mol. Cell Biol. 2007;8:379–393. - PubMed
-
- Peters J.M. The anaphase promoting complex/cyclosome: A machine designed to destroy. Nat. Rev. Mol. Cell Biol. 2006;7:644–656. - PubMed
-
- Pinsky B.A., Biggins S. The spindle checkpoint: Tension versus attachment. Trends Cell Biol. 2005;15:486–493. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
