

Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150 Glued and late endosome positioning
- PMID: 19564404
- PMCID: PMC2712958
- DOI: 10.1083/jcb.200811005
Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150 Glued and late endosome positioning
Abstract
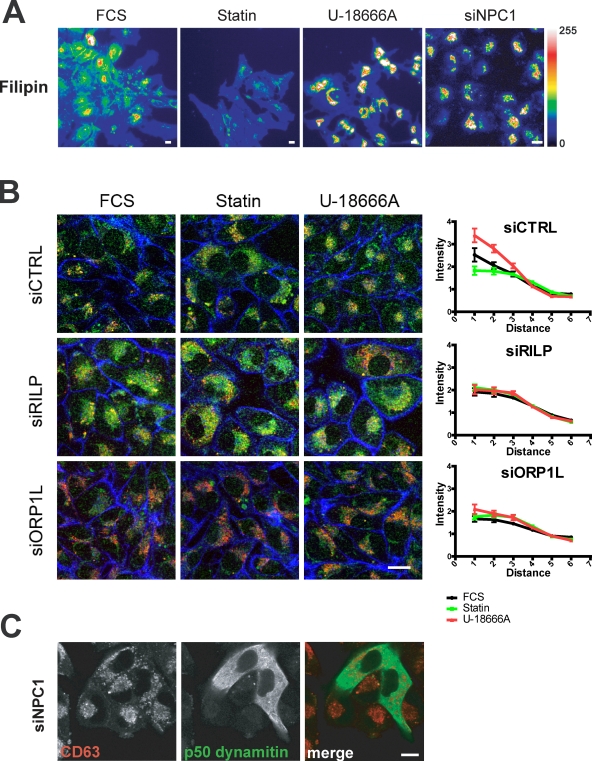
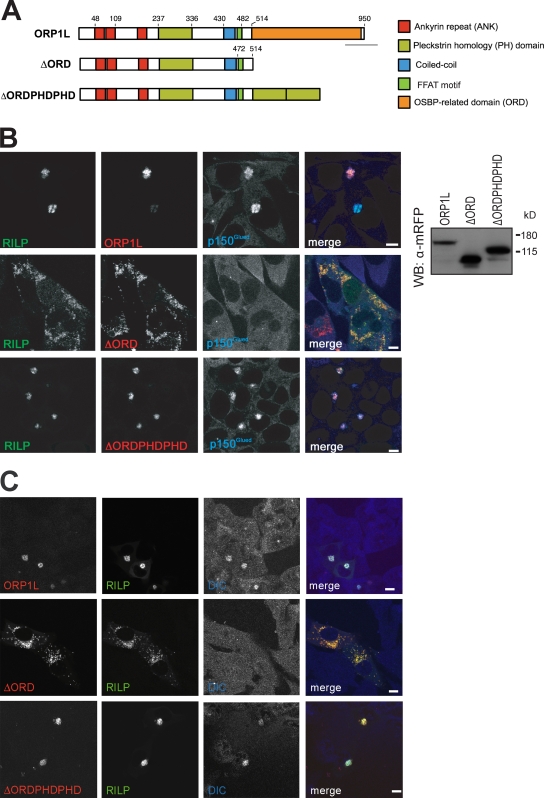
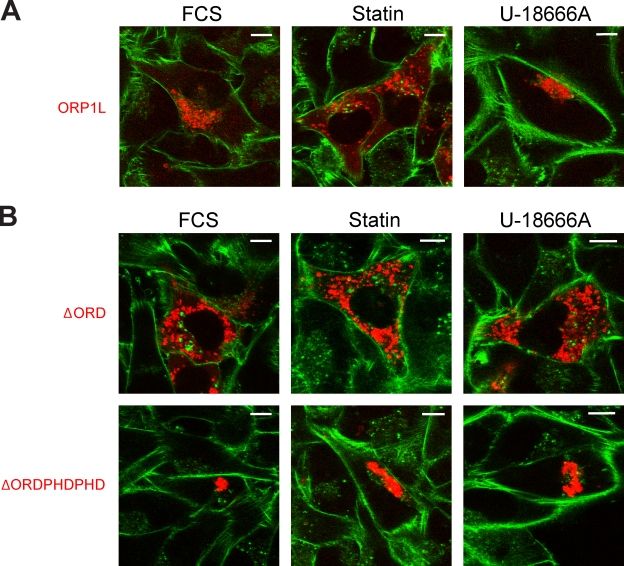
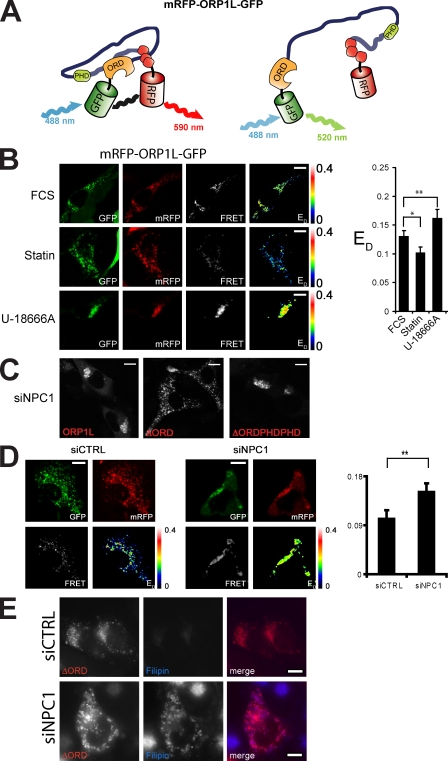
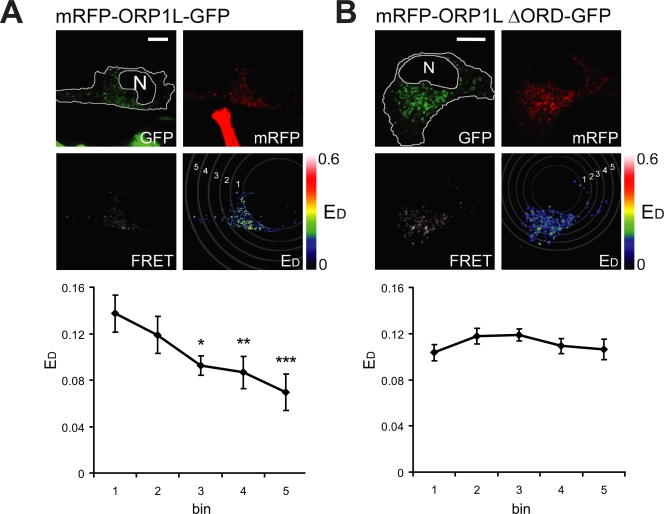
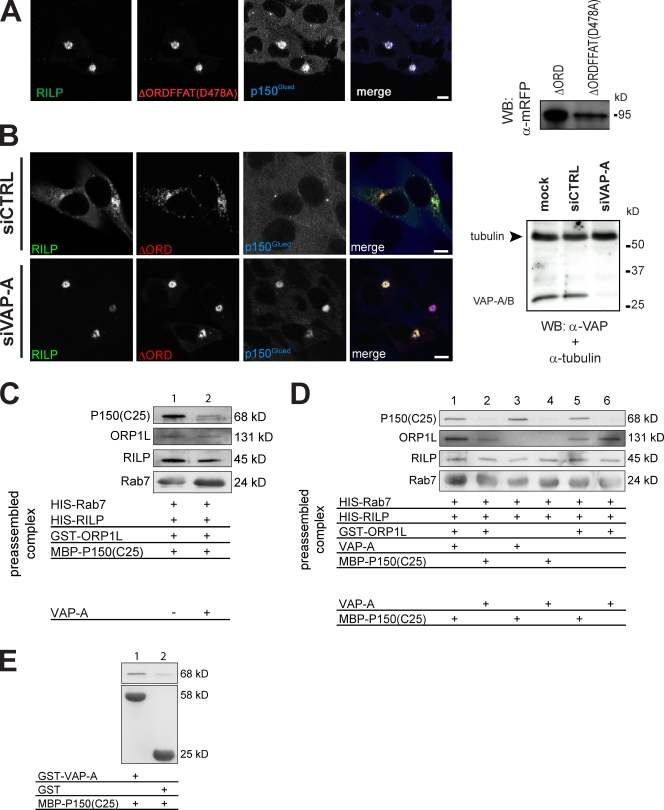
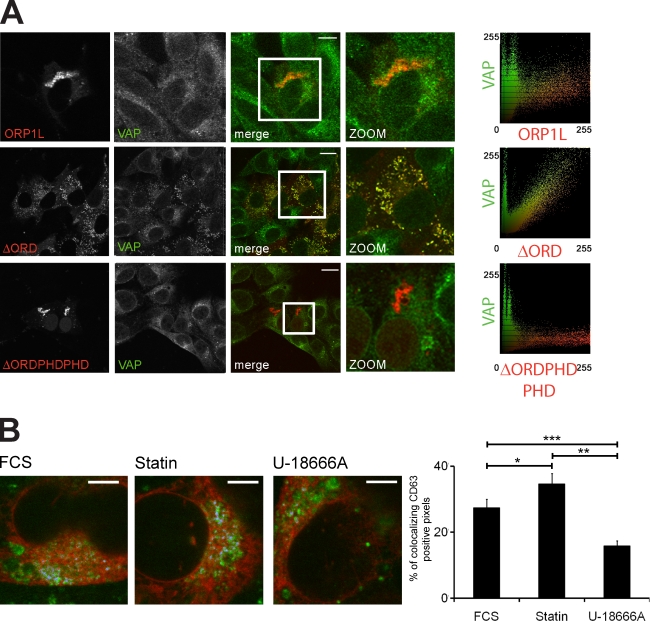
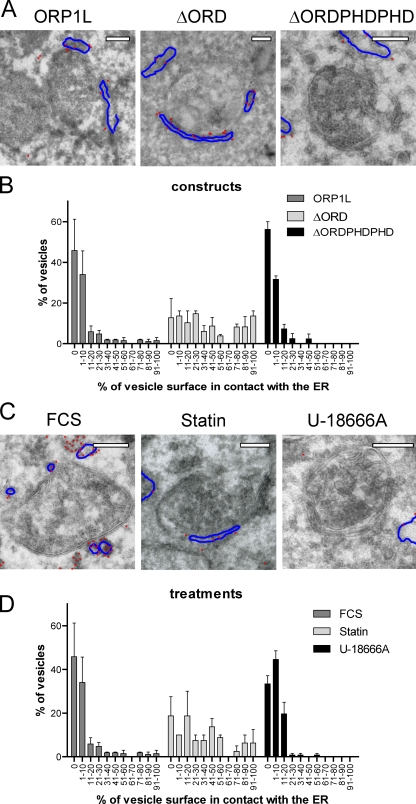
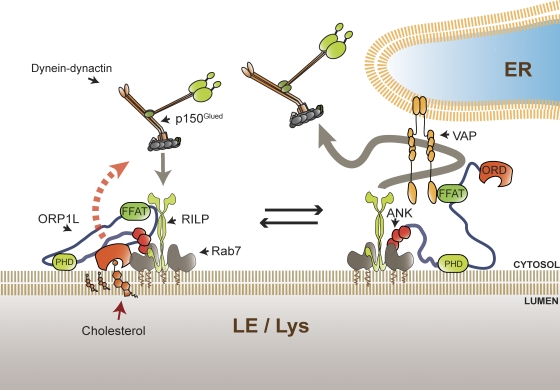
Late endosomes (LEs) have characteristic intracellular distributions determined by their interactions with various motor proteins. Motor proteins associated to the dynactin subunit p150(Glued) bind to LEs via the Rab7 effector Rab7-interacting lysosomal protein (RILP) in association with the oxysterol-binding protein ORP1L. We found that cholesterol levels in LEs are sensed by ORP1L and are lower in peripheral vesicles. Under low cholesterol conditions, ORP1L conformation induces the formation of endoplasmic reticulum (ER)-LE membrane contact sites. At these sites, the ER protein VAP (VAMP [vesicle-associated membrane protein]-associated ER protein) can interact in trans with the Rab7-RILP complex to remove p150(Glued) and associated motors. LEs then move to the microtubule plus end. Under high cholesterol conditions, as in Niemann-Pick type C disease, this process is prevented, and LEs accumulate at the microtubule minus end as the result of dynein motor activity. These data explain how the ER and cholesterol control the association of LEs with motor proteins and their positioning in cells.
Figures
Similar articles
-
Activation of endosomal dynein motors by stepwise assembly of Rab7-RILP-p150Glued, ORP1L, and the receptor betalll spectrin.J Cell Biol. 2007 Feb 12;176(4):459-71. doi: 10.1083/jcb.200606077. Epub 2007 Feb 5. J Cell Biol. 2007. PMID: 17283181 Free PMC article.
-
Late endosomal transport and tethering are coupled processes controlled by RILP and the cholesterol sensor ORP1L.J Cell Sci. 2013 Aug 1;126(Pt 15):3462-74. doi: 10.1242/jcs.129270. Epub 2013 May 31. J Cell Sci. 2013. PMID: 23729732
-
RILP Induces Cholesterol Accumulation in Lysosomes by Inhibiting Endoplasmic Reticulum-Endolysosome Interactions.Cells. 2024 Aug 6;13(16):1313. doi: 10.3390/cells13161313. Cells. 2024. PMID: 39195203 Free PMC article.
-
ER-endosome contact sites in endosome positioning and protrusion outgrowth.Biochem Soc Trans. 2016 Apr 15;44(2):441-6. doi: 10.1042/BST20150246. Biochem Soc Trans. 2016. PMID: 27068952 Review.
-
Touché! STARD3 and STARD3NL tether the ER to endosomes.Biochem Soc Trans. 2016 Apr 15;44(2):493-8. doi: 10.1042/BST20150269. Biochem Soc Trans. 2016. PMID: 27068960 Review.
Cited by
-
circTP63 promotes prostate cancer progression via miR-421/VAMP associated protein A axis.J Cancer. 2024 Aug 19;15(16):5451-5461. doi: 10.7150/jca.99561. eCollection 2024. J Cancer. 2024. PMID: 39247600 Free PMC article.
-
Sphingosine kinases regulate ER contacts with late endocytic organelles and cholesterol trafficking.Proc Natl Acad Sci U S A. 2022 Sep 27;119(39):e2204396119. doi: 10.1073/pnas.2204396119. Epub 2022 Sep 19. Proc Natl Acad Sci U S A. 2022. PMID: 36122218 Free PMC article.
-
The endolysosomal system in conventional and unconventional protein secretion.J Cell Biol. 2024 Sep 2;223(9):e202404152. doi: 10.1083/jcb.202404152. Epub 2024 Aug 12. J Cell Biol. 2024. PMID: 39133205 Free PMC article. Review.
-
DENND6A links Arl8b to a Rab34/RILP/dynein complex, regulating lysosomal positioning and autophagy.Nat Commun. 2024 Jan 31;15(1):919. doi: 10.1038/s41467-024-44957-1. Nat Commun. 2024. PMID: 38296963 Free PMC article.
-
Endoplasmic reticulum-endosome contact increases as endosomes traffic and mature.Mol Biol Cell. 2013 Apr;24(7):1030-40. doi: 10.1091/mbc.E12-10-0733. Epub 2013 Feb 6. Mol Biol Cell. 2013. PMID: 23389631 Free PMC article.
References
-
- Bastiaens P.I., Squire A. 1999. Fluorescence lifetime imaging microscopy: spatial resolution of biochemical processes in the cell.Trends Cell Biol. 9:48–52 - PubMed
-
- Brown C.L., Maier K.C., Stauber T., Ginkel L.M., Wordeman L., Vernos I., Schroer T.A. 2005. Kinesin-2 is a motor for late endosomes and lysosomes.Traffic. 6:1114–1124 - PubMed
-
- Calafat J., Janssen H., Stahle-Backdahl M., Zuurbier A.E., Knol E.F., Egesten A. 1997. Human monocytes and neutrophils store transforming growth factor-alpha in a subpopulation of cytoplasmic granules.Blood. 90:1255–1266 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
