

3' cis-acting elements that contribute to the competence and efficiency of Japanese encephalitis virus genome replication: functional importance of sequence duplications, deletions, and substitutions
- PMID: 19494005
- PMCID: PMC2715749
- DOI: 10.1128/JVI.02541-08
3' cis-acting elements that contribute to the competence and efficiency of Japanese encephalitis virus genome replication: functional importance of sequence duplications, deletions, and substitutions
Abstract
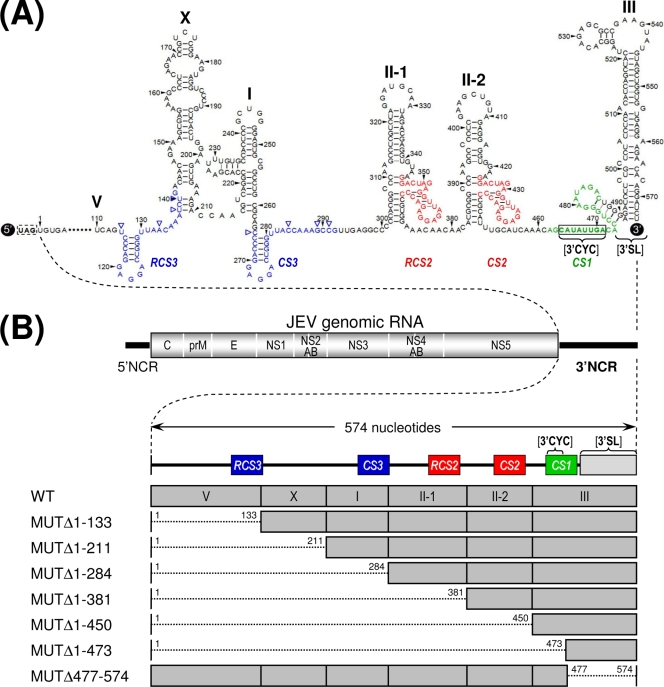
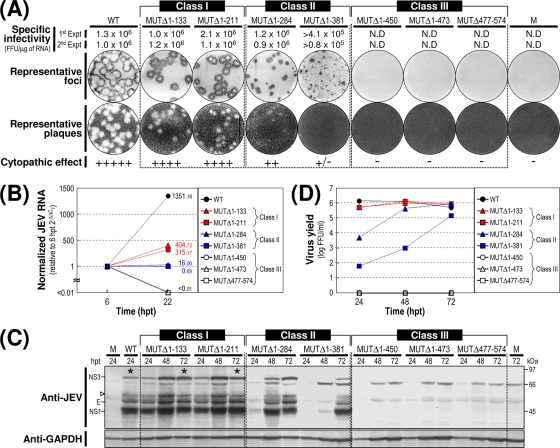
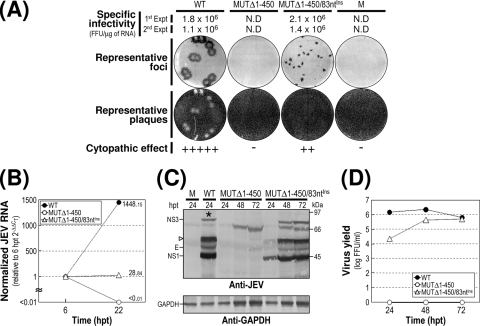
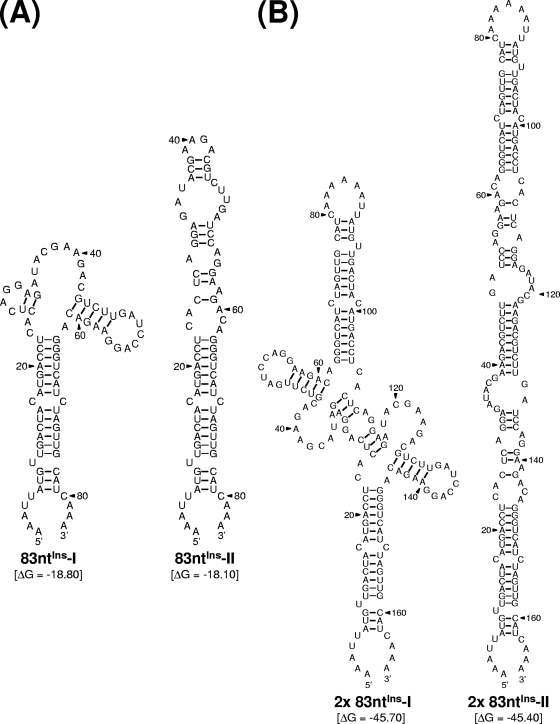
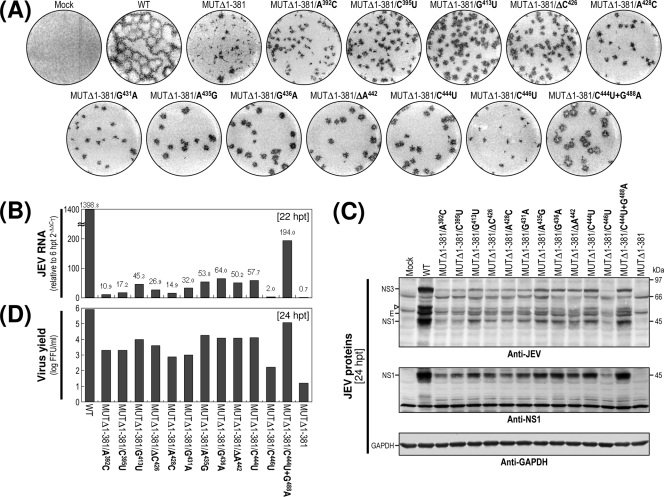
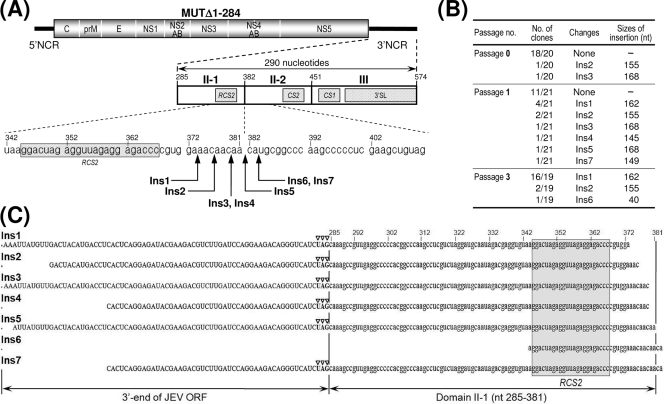
The positive-strand RNA genome of Japanese encephalitis virus (JEV) terminates in a highly conserved 3'-noncoding region (3'NCR) of six domains (V, X, I, II-1, II-2, and III in the 5'-to-3' direction). By manipulating the JEV genomic RNA, we have identified important roles for RNA elements present within the 574-nucleotide 3'NCR in viral replication. The two 3'-proximal domains (II-2 and III) were sufficient for RNA replication and virus production, whereas the remaining four (V, X, I, and II-1) were dispensable for RNA replication competence but required for maximal replication efficiency. Surprisingly, a lethal mutant lacking all of the 3'NCR except domain III regained viability through pseudoreversion by duplicating an 83-nucleotide sequence from the 3'-terminal region of the viral open reading frame. Also, two viable mutants displayed severe genetic instability; these two mutants rapidly developed 12 point mutations in domain II-2 in the mutant lacking domains V, X, I, and II-1 and showed the duplication of seven upstream sequences of various sizes at the junction between domains II-1 and II-2 in the mutant lacking domains V, X, and I. In all cases, the introduction of these spontaneous mutations led to an increase in RNA production that paralleled the level of protein accumulation and virus yield. Interestingly, the mutant lacking domains V, X, I, and II-1 was able to replicate in hamster BHK-21 and human neuroblastoma SH-SY5Y cells but not in mosquito C6/36 cells, indicating a cell type-specific restriction of its viral replication. Thus, our findings provide the basis for a detailed map of the 3' cis-acting elements in JEV genomic RNA, which play an essential role in viral replication. They also provide experimental evidence for the function of 3' direct repeat sequences and suggest possible mechanisms for the emergence of these sequences in the 3'NCR of JEV and perhaps in other flaviviruses.
Figures
Similar articles
-
Sequence duplication in 3' UTR modulates virus replication and virulence of Japanese encephalitis virus.Emerg Microbes Infect. 2022 Dec;11(1):123-135. doi: 10.1080/22221751.2021.2016354. Emerg Microbes Infect. 2022. PMID: 34877923 Free PMC article.
-
Inhibition of Japanese encephalitis virus replication by peptide nucleic acids targeting cis-acting elements on the plus- and minus-strands of viral RNA.Antiviral Res. 2009 Jun;82(3):122-33. doi: 10.1016/j.antiviral.2009.02.187. Epub 2009 Feb 20. Antiviral Res. 2009. PMID: 19428603
-
Genetic variation in the 3' non-coding region of dengue viruses.Virology. 2001 Mar 1;281(1):75-87. doi: 10.1006/viro.2000.0748. Virology. 2001. PMID: 11222098
-
Genome cyclization as strategy for flavivirus RNA replication.Virus Res. 2009 Feb;139(2):230-9. doi: 10.1016/j.virusres.2008.07.016. Epub 2008 Sep 9. Virus Res. 2009. PMID: 18703097 Free PMC article. Review.
-
Cis-acting RNA elements in human and animal plus-strand RNA viruses.Biochim Biophys Acta. 2009 Sep-Oct;1789(9-10):495-517. doi: 10.1016/j.bbagrm.2009.09.007. Epub 2009 Sep 23. Biochim Biophys Acta. 2009. PMID: 19781674 Free PMC article. Review.
Cited by
-
Profiling of viral proteins expressed from the genomic RNA of Japanese encephalitis virus using a panel of 15 region-specific polyclonal rabbit antisera: implications for viral gene expression.PLoS One. 2015 Apr 27;10(4):e0124318. doi: 10.1371/journal.pone.0124318. eCollection 2015. PLoS One. 2015. PMID: 25915765 Free PMC article.
-
Japanese encephalitis: the virus and vaccines.Hum Vaccin Immunother. 2014;10(2):263-79. doi: 10.4161/hv.26902. Epub 2013 Oct 25. Hum Vaccin Immunother. 2014. PMID: 24161909 Free PMC article. Review.
-
A molecularly cloned, live-attenuated japanese encephalitis vaccine SA14-14-2 virus: a conserved single amino acid in the ij Hairpin of the Viral E glycoprotein determines neurovirulence in mice.PLoS Pathog. 2014 Jul 31;10(7):e1004290. doi: 10.1371/journal.ppat.1004290. eCollection 2014 Jul. PLoS Pathog. 2014. PMID: 25077483 Free PMC article.
-
Zika virus: An emerging flavivirus.J Microbiol. 2017 Mar;55(3):204-219. doi: 10.1007/s12275-017-7063-6. Epub 2017 Feb 28. J Microbiol. 2017. PMID: 28243937 Review.
-
Nonsuppurative encephalomyelitis in a calf in Japan and isolation of Japanese encephalitis virus genotype 1 from the affected calf.J Clin Microbiol. 2013 Oct;51(10):3448-53. doi: 10.1128/JCM.00737-13. Epub 2013 Jul 24. J Clin Microbiol. 2013. PMID: 23885004 Free PMC article.
References
-
- Ackermann, M., and R. Padmanabhan. 2001. De novo synthesis of RNA by the dengue virus RNA-dependent RNA polymerase exhibits temperature dependence at the initiation but not elongation phase. J. Biol. Chem. 27639926-39937. - PubMed
-
- Alvarez, D. E., A. L. De Lella Ezcurra, S. Fucito, and A. V. Gamarnik. 2005a. Role of RNA structures present at the 3′UTR of dengue virus on translation, RNA synthesis, and viral replication. Virology 339200-212. - PubMed
-
- Alvarez, D. E., C. V. Filomatori, and A. V. Gamarnik. 2008. Functional analysis of dengue virus cyclization sequences located at the 5′ and 3′UTRs. Virology 375223-235. - PubMed
-
- Ball, L. A. 1997. Nodavirus RNA recombination. Semin. Virol. 895-100.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
