

Broadly neutralizing anti-HIV-1 antibodies disrupt a hinge-related function of gp41 at the membrane interface
- PMID: 19458040
- PMCID: PMC2690059
- DOI: 10.1073/pnas.0901474106
Broadly neutralizing anti-HIV-1 antibodies disrupt a hinge-related function of gp41 at the membrane interface
Abstract
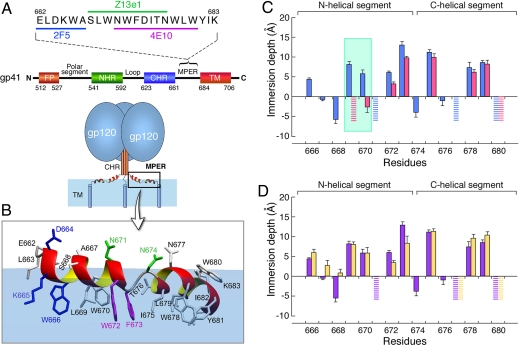
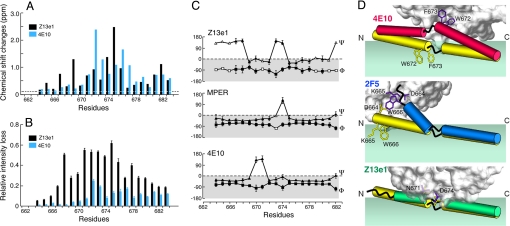
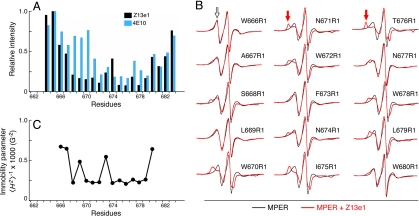
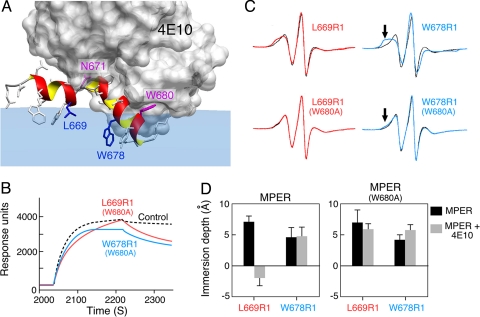
A vaccine capable of stimulating protective antiviral antibody responses is needed to curtail the global AIDS epidemic caused by HIV-1. Although rarely elicited during the course of natural infection or upon conventional vaccination, the membrane-proximal ectodomain region (MPER) of the HIV-1 glycoprotein of M(r) 41,000 (gp41) envelope protein subunit is the target of 3 such human broadly neutralizing antibodies (BNAbs): 4E10, 2F5, and Z13e1. How these BNAbs bind to their lipid-embedded epitopes and mediate antiviral activity is unclear, but such information might offer important insight into a worldwide health imperative. Here, EPR and NMR techniques were used to define the manner in which these BNAbs differentially recognize viral membrane-encrypted residues configured within the L-shaped helix-hinge-helix MPER segment. Two distinct modes of antibody-mediated interference of viral infection were identified. 2F5, like 4E10, induces large conformational changes in the MPER relative to the membrane. However, although 4E10 straddles the hinge and extracts residues W672 and F673, 2F5 lifts up residues N-terminal to the hinge region, exposing L669 and W670. In contrast, Z13e1 effects little change in membrane orientation or conformation, but rather immobilizes the MPER hinge through extensive rigidifying surface contacts. Thus, BNAbs disrupt HIV-1 MPER fusogenic functions critical for virus entry into human CD4 T cells and macrophages either by preventing hinge motion or by perturbing MPER orientation. HIV-1 MPER features important for targeted vaccine design have been revealed, the implications of which extend to BNAb targets on other viral fusion proteins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
HIV-1 broadly neutralizing antibody extracts its epitope from a kinked gp41 ectodomain region on the viral membrane.Immunity. 2008 Jan;28(1):52-63. doi: 10.1016/j.immuni.2007.11.018. Immunity. 2008. PMID: 18191596
-
Anti-human immunodeficiency virus type 1 (HIV-1) antibodies 2F5 and 4E10 require surprisingly few crucial residues in the membrane-proximal external region of glycoprotein gp41 to neutralize HIV-1.J Virol. 2005 Jan;79(2):1252-61. doi: 10.1128/JVI.79.2.1252-1261.2005. J Virol. 2005. PMID: 15613352 Free PMC article.
-
The broadly neutralizing anti-human immunodeficiency virus type 1 4E10 monoclonal antibody is better adapted to membrane-bound epitope recognition and blocking than 2F5.J Virol. 2008 Sep;82(18):8986-96. doi: 10.1128/JVI.00846-08. Epub 2008 Jul 2. J Virol. 2008. PMID: 18596094 Free PMC article.
-
Antigp41 membrane proximal external region antibodies and the art of using the membrane for neutralization.Curr Opin HIV AIDS. 2017 May;12(3):250-256. doi: 10.1097/COH.0000000000000364. Curr Opin HIV AIDS. 2017. PMID: 28422789 Review.
-
Neutralizing Antibodies Targeting HIV-1 gp41.Viruses. 2020 Oct 23;12(11):1210. doi: 10.3390/v12111210. Viruses. 2020. PMID: 33114242 Free PMC article. Review.
Cited by
-
Influences on trimerization and aggregation of soluble, cleaved HIV-1 SOSIP envelope glycoprotein.J Virol. 2013 Sep;87(17):9873-85. doi: 10.1128/JVI.01226-13. Epub 2013 Jul 3. J Virol. 2013. PMID: 23824824 Free PMC article.
-
Association of enhanced HIV-1 neutralization by a single Y681H substitution in gp41 with increased gp120-CD4 interaction and macrophage infectivity.PLoS One. 2012;7(5):e37157. doi: 10.1371/journal.pone.0037157. Epub 2012 May 14. PLoS One. 2012. PMID: 22606344 Free PMC article.
-
Aromatic residues at the edge of the antibody combining site facilitate viral glycoprotein recognition through membrane interactions.Proc Natl Acad Sci U S A. 2010 Jan 26;107(4):1529-34. doi: 10.1073/pnas.0909680107. Epub 2010 Jan 4. Proc Natl Acad Sci U S A. 2010. PMID: 20080706 Free PMC article.
-
Immunogenicity of membrane-bound HIV-1 gp41 membrane-proximal external region (MPER) segments is dominated by residue accessibility and modulated by stereochemistry.J Biol Chem. 2013 Nov 1;288(44):31888-901. doi: 10.1074/jbc.M113.494609. Epub 2013 Sep 18. J Biol Chem. 2013. PMID: 24047898 Free PMC article.
-
Structure-based design of a protein immunogen that displays an HIV-1 gp41 neutralizing epitope.J Mol Biol. 2011 Dec 2;414(3):460-76. doi: 10.1016/j.jmb.2011.10.014. Epub 2011 Oct 15. J Mol Biol. 2011. PMID: 22033480 Free PMC article.
References
-
- Feng Y, Broder CC, Kennedy PE, Berger EA. HIV-1 entry cofactor: Functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science. 1996;272:872–877. - PubMed
-
- Chan DC, Fass D, Berger JM, Kim PS. Core structure of gp41 from the HIV envelope glycoprotein. Cell. 1997;89:263–273. - PubMed
-
- Wei X, et al. Antibody neutralization and escape by HIV-1. Nature. 2003;422:307–312. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
