

Regulation of luminal acidification in the male reproductive tract via cell-cell crosstalk
- PMID: 19448084
- PMCID: PMC2683015
- DOI: 10.1242/jeb.027284
Regulation of luminal acidification in the male reproductive tract via cell-cell crosstalk
Abstract
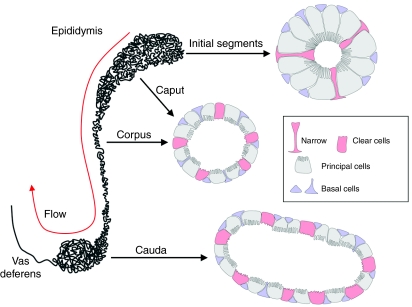
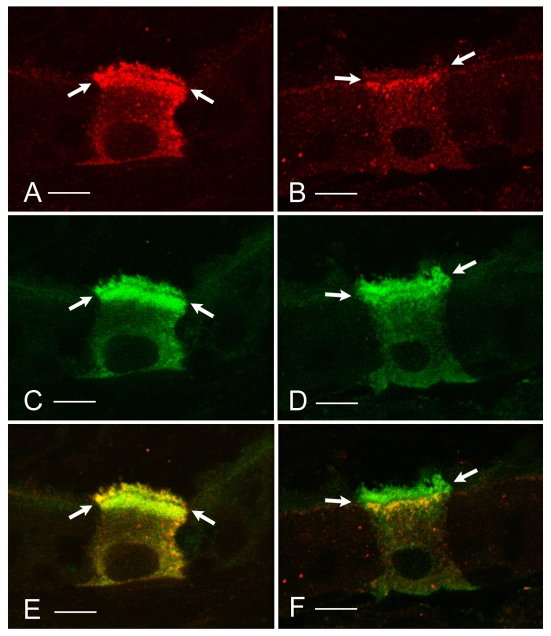
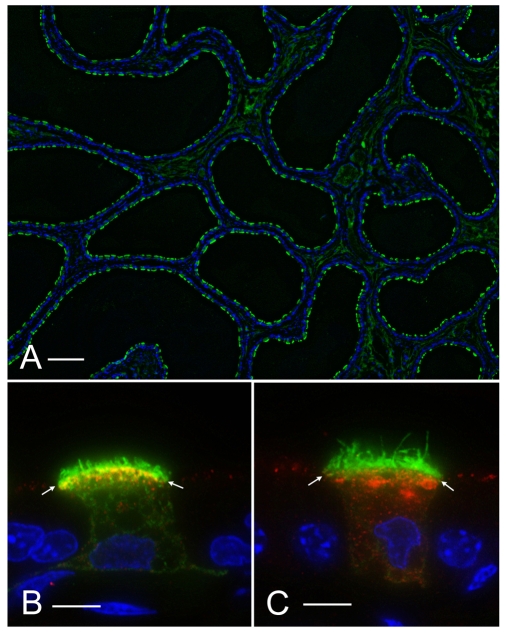
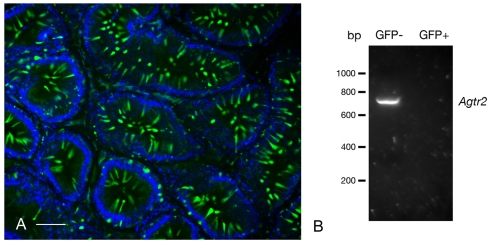
In the epididymis, spermatozoa acquire their ability to become motile and to fertilize an egg. A luminal acidic pH and a low bicarbonate concentration help keep spermatozoa in a quiescent state during their maturation and storage in this organ. Net proton secretion is crucial to maintain the acidity of the luminal fluid in the epididymis. A sub-population of epithelial cells, the clear cells, express high levels of the proton-pumping V-ATPase in their apical membrane and are important contributors to luminal acidification. This review describes selected aspects of V-ATPase regulation in clear cells. The assembly of a particular set of V-ATPase subunit isoforms governs the targeting of the pump to the apical plasma membrane. Regulation of V-ATPase-dependent proton secretion occurs via recycling mechanisms. The bicarbonate-activated adenylyl cyclase is involved in the non-hormonal regulation of V-ATPase recycling, following activation of bicarbonate secretion by principal cells. The V-ATPase is also regulated in a paracrine manner by luminal angiotensin II by activation of the angiotensin II type 2 receptor (AGTR2), which is located in basal cells. Basal cells have the remarkable property of extending long and slender cytoplasmic projections that cross the tight junction barrier to monitor the luminal environment. Clear cells are activated by a nitric oxide signal that originates from basal cells. Thus, a complex interplay between the different cell types present in the epithelium leads to activation of the luminal acidifying capacity of the epididymis, a process that is crucial for sperm maturation and storage.
Figures







Similar articles
-
Establishment of cell-cell cross talk in the epididymis: control of luminal acidification.J Androl. 2011 Nov-Dec;32(6):576-86. doi: 10.2164/jandrol.111.012971. Epub 2011 Mar 25. J Androl. 2011. PMID: 21441423 Free PMC article. Review.
-
Unravelling purinergic regulation in the epididymis: activation of V-ATPase-dependent acidification by luminal ATP and adenosine.J Physiol. 2019 Apr;597(7):1957-1973. doi: 10.1113/JP277565. Epub 2019 Feb 28. J Physiol. 2019. PMID: 30746715 Free PMC article.
-
Modulation of the actin cytoskeleton via gelsolin regulates vacuolar H+-ATPase recycling.J Biol Chem. 2005 Mar 4;280(9):8452-63. doi: 10.1074/jbc.M412750200. Epub 2004 Dec 9. J Biol Chem. 2005. PMID: 15591047
-
The Involvement of the Chemokine RANTES in Regulating Luminal Acidification in Rat Epididymis.Front Immunol. 2020 Sep 25;11:583274. doi: 10.3389/fimmu.2020.583274. eCollection 2020. Front Immunol. 2020. PMID: 33072131 Free PMC article.
-
Regulation of vacuolar proton pumping ATPase-dependent luminal acidification in the epididymis.Asian J Androl. 2007 Jul;9(4):476-82. doi: 10.1111/j.1745-7262.2007.00299.x. Asian J Androl. 2007. PMID: 17589784 Review.
Cited by
-
Pulmonary Arterial Hypertension Affects Sperm Quality and Epididymis Function in Sedentary and Exercised Wistar Rats.Reprod Sci. 2024 Nov;31(11):3485-3497. doi: 10.1007/s43032-024-01713-6. Epub 2024 Oct 2. Reprod Sci. 2024. PMID: 39356456
-
The significance of single-cell transcriptome analysis in epididymis research.Front Cell Dev Biol. 2024 Mar 21;12:1357370. doi: 10.3389/fcell.2024.1357370. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 38577504 Free PMC article.
-
Recent advances in acid sensing by G protein coupled receptors.Pflugers Arch. 2024 Apr;476(4):445-455. doi: 10.1007/s00424-024-02919-y. Epub 2024 Feb 10. Pflugers Arch. 2024. PMID: 38340167 Free PMC article. Review.
-
Effects of Heparan sulfate acetyl-CoA: Alpha-glucosaminide N-acetyltransferase (HGSNAT) inactivation on the structure and function of epithelial and immune cells of the testis and epididymis and sperm parameters in adult mice.PLoS One. 2023 Sep 27;18(9):e0292157. doi: 10.1371/journal.pone.0292157. eCollection 2023. PLoS One. 2023. PMID: 37756356 Free PMC article.
-
Differential expression and localization of tight junction proteins in the goat epididymis.J Anim Sci Technol. 2022 May;64(3):500-514. doi: 10.5187/jast.2022.e13. Epub 2022 May 31. J Anim Sci Technol. 2022. PMID: 35709128 Free PMC article.
References
-
- Acott, T. S. and Carr, D. W. (1984). Inhibition of bovine spermatozoa by caudal epididymal fluid: II. Interaction of pH and a quiescence factor. Biol. Reprod. 30, 926-935. - PubMed
-
- Bagnis, C., Marsolais, M., Biemesderfer, D., Laprade, R. and Breton, S. (2001). Na(+)/H(+)-exchange activity and immunolocalization of NHE3 in rat epididymis. Am. J. Physiol. Renal Physiol. 280, F426-F436. - PubMed
-
- Beaulieu, V., Da Silva, N., Pastor-Soler, N., Brown, C. R., Smith, P. J., Brown, D. and Breton, S. (2005). Modulation of the actin cytoskeleton via gelsolin regulates vacuolar H+ATPase (V-ATPase) recycling. J. Biol. Chem. 280, 8452-8463. - PubMed
-
- Beyenbach, K. W. and Wieczorek, H. (2006). The V-type H+ ATPase: molecular structure and function, physiological roles and regulation. J. Exp. Biol. 209, 577-589. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
