

Sexual development in Lucilia cuprina (Diptera, Calliphoridae) is controlled by the transformer gene
- PMID: 19433631
- PMCID: PMC2710159
- DOI: 10.1534/genetics.109.100982
Sexual development in Lucilia cuprina (Diptera, Calliphoridae) is controlled by the transformer gene
Abstract
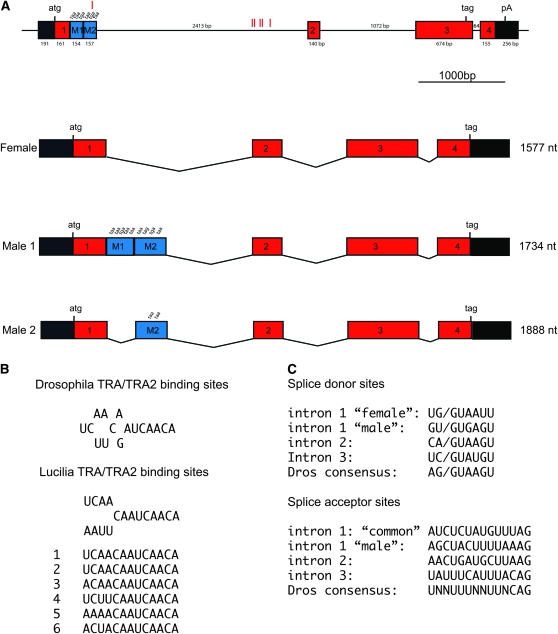
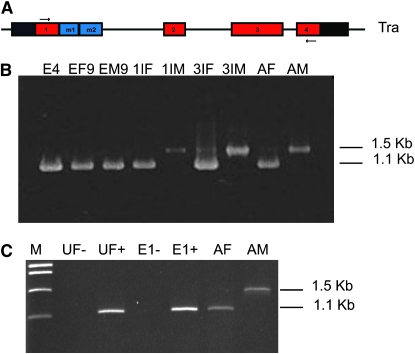
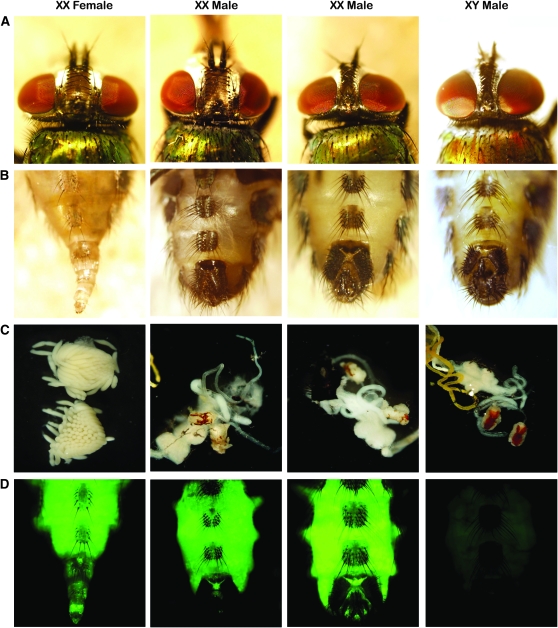
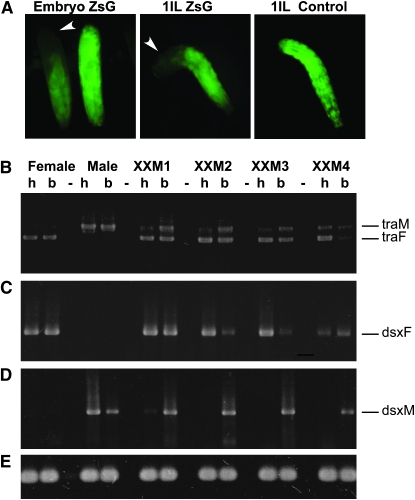
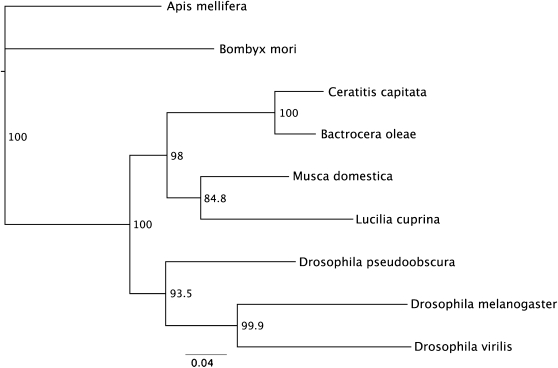
Insects use an amazing variety of genetic systems to control sexual development. A Y-linked male determining gene (M) controls sex in the Australian sheep blowfly Lucilia cuprina, an important pest insect. In this study, we isolated the L. cuprina transformer (Lctra) and transformer2 (Lctra2) genes, which are potential targets of M. The LCTRA and LCTRA2 proteins are significantly more similar to homologs from tephritid insects than Drosophila. The Lctra transcript is alternatively spliced such that only females make a full-length protein and the presence of six TRA/TRA2 binding sites in the female first intron suggest that Lctra splicing is autoregulated as in tephritids. LCTRA is essential for female development as RNAi knockdown of Lctra mRNA leads to the development of male genitalia in XX adults. Analysis of Lctra expression during development shows that early and midstage male and female embryos express the female form of Lctra and males express only the male form by the first instar larval stage. Our results suggest that an autoregulatory loop sustains female development and that expression of M inhibits Lctra autoregulation, switching its splicing to the male form. The conservation of tra function and regulation in a Calliphorid insect shows that this sex determination system is not confined to Tephritidae. Isolation of these genes is an important step toward the development of a strain of L. cuprina suitable for a genetic control program.
Figures


Similar articles
-
Conservation and sex-specific splicing of the doublesex gene in the economically important pest species Lucilia cuprina.J Genet. 2010 Sep;89(3):279-85. doi: 10.1007/s12041-010-0039-5. J Genet. 2010. PMID: 20876994
-
Conditional knockdown of transformer in sheep blow fly suggests a role in repression of dosage compensation and potential for population suppression.PLoS Genet. 2021 Oct 18;17(10):e1009792. doi: 10.1371/journal.pgen.1009792. eCollection 2021 Oct. PLoS Genet. 2021. PMID: 34662332 Free PMC article.
-
Conservation and sex-specific splicing of the transformer gene in the calliphorids Cochliomyia hominivorax, Cochliomyia macellaria and Lucilia sericata.PLoS One. 2013;8(2):e56303. doi: 10.1371/journal.pone.0056303. Epub 2013 Feb 7. PLoS One. 2013. PMID: 23409170 Free PMC article.
-
Sex determination mechanisms in the Calliphoridae (blow flies).Sex Dev. 2014;8(1-3):29-37. doi: 10.1159/000357132. Epub 2013 Dec 21. Sex Dev. 2014. PMID: 24401179 Review.
-
Development and evaluation of male-only strains of the Australian sheep blowfly, Lucilia cuprina.BMC Genet. 2014;15 Suppl 2(Suppl 2):S3. doi: 10.1186/1471-2156-15-S2-S3. Epub 2014 Dec 1. BMC Genet. 2014. PMID: 25472415 Free PMC article. Review.
Cited by
-
The Sex Determination Cascade in the Silkworm.Genes (Basel). 2021 Feb 23;12(2):315. doi: 10.3390/genes12020315. Genes (Basel). 2021. PMID: 33672402 Free PMC article. Review.
-
Genome sequence of Phormia regina Meigen (Diptera: Calliphoridae): implications for medical, veterinary and forensic research.BMC Genomics. 2016 Oct 28;17(1):842. doi: 10.1186/s12864-016-3187-z. BMC Genomics. 2016. PMID: 27793085 Free PMC article.
-
The role of the Drosophila LAMMER protein kinase DOA in somatic sex determination.J Genet. 2010 Sep;89(3):271-7. doi: 10.1007/s12041-010-0038-6. J Genet. 2010. PMID: 20876993
-
The gene transformer-2 of Sciara (Diptera, Nematocera) and its effect on Drosophila sexual development.BMC Dev Biol. 2011 Mar 15;11:19. doi: 10.1186/1471-213X-11-19. BMC Dev Biol. 2011. PMID: 21406087 Free PMC article.
-
Genomic organization and splicing evolution of the doublesex gene, a Drosophila regulator of sexual differentiation, in the dengue and yellow fever mosquito Aedes aegypti.BMC Evol Biol. 2011 Feb 10;11:41. doi: 10.1186/1471-2148-11-41. BMC Evol Biol. 2011. PMID: 21310052 Free PMC article.
References
-
- Beck, T., B. Moir and T. Meppem, 1985. The cost of parasites to the Australian sheep industry. Q. Rev. Rural Econ. 7 336–343.
-
- Bedo, D. G., and G. G. Foster, 1985. Cytogenetic mapping of the male-determining region of Lucilia cuprina (Diptera: Calliphoridae). Chromosoma 92 344–350.
-
- Brogna, S., P. V. Benos, G. Gasperi and C. Savakis, 2001. The Drosophila alcohol dehydrogenase gene may have evolved independently of the functionally homologous medfly, olive fly, and flesh fly genes. Mol. Biol. Evol. 18 322–329. - PubMed
-
- Burghardt, G., M. Hediger, C. Siegenthaler, M. Moser, A. Dübendorfer et al., 2005. The transformer2 gene in Musca domestica is required for selecting and maintaining the female pathway of development. Dev. Genes Evol. 215 165–176. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
