

Prediction of novel families of enzymes involved in oxidative and other complex modifications of bases in nucleic acids
- PMID: 19411852
- PMCID: PMC2995806
- DOI: 10.4161/cc.8.11.8580
Prediction of novel families of enzymes involved in oxidative and other complex modifications of bases in nucleic acids
Abstract
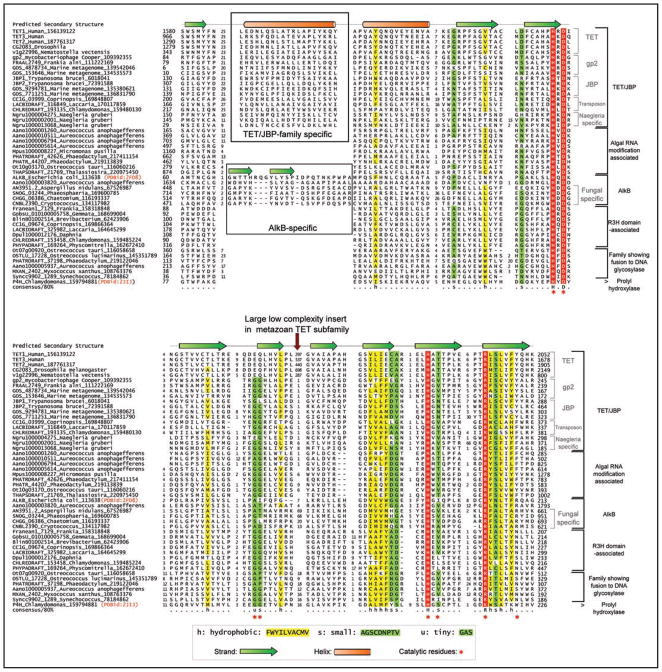
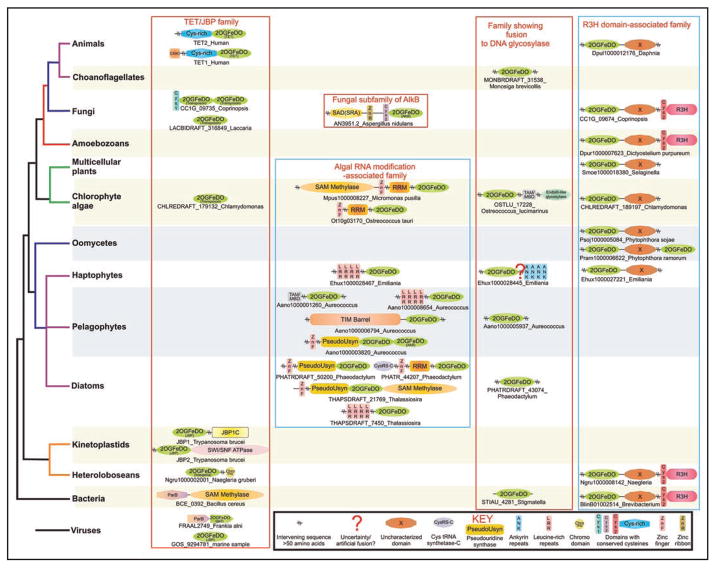
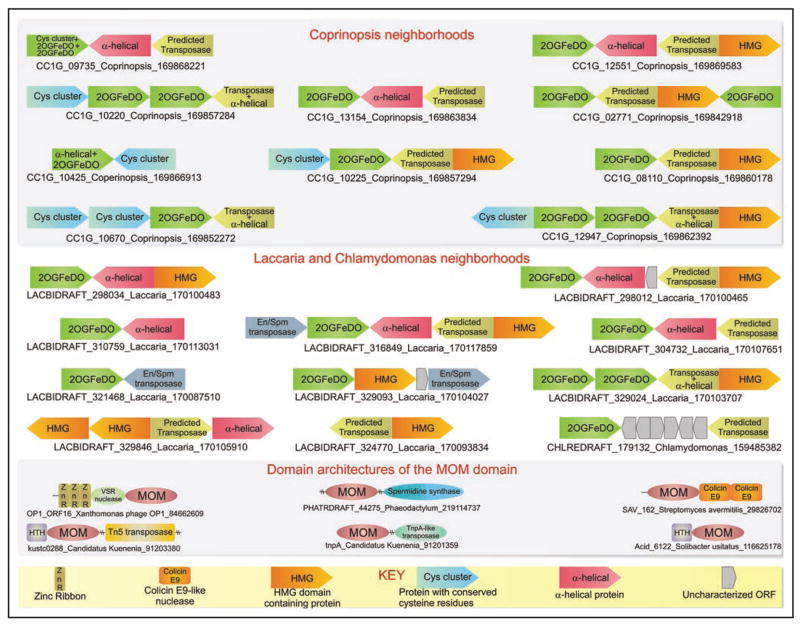
Modified bases in nucleic acids present a layer of information that directs biological function over and beyond the coding capacity of the conventional bases. While a large number of modified bases have been identified, many of the enzymes generating them still remain to be discovered. Recently, members of the 2-oxoglutarate- and iron(II)-dependent dioxygenase super-family, which modify diverse substrates from small molecules to biopolymers, were predicted and subsequently confirmed to catalyze oxidative modification of bases in nucleic acids. Of these, two distinct families, namely the AlkB and the kinetoplastid base J binding proteins (JBP) catalyze in situ hydroxylation of bases in nucleic acids. Using sensitive computational analysis of sequences, structures and contextual information from genomic structure and protein domain architectures, we report five distinct families of 2-oxoglutarate- and iron(II)-dependent dioxygenase that we predict to be involved in nucleic acid modifications. Among the DNA-modifying families, we show that the dioxygenase domains of the kinetoplastid base J-binding proteins belong to a larger family that includes the Tet proteins, prototyped by the human oncogene Tet1, and proteins from basidiomycete fungi, chlorophyte algae, heterolobosean amoeboflagellates and bacteriophages. We present evidence that some of these proteins are likely to be involved in oxidative modification of the 5-methyl group of cytosine leading to the formation of 5-hydroxymethylcytosine. The Tet/JBP homologs from basidiomycete fungi such as Laccaria and Coprinopsis show large lineage-specific expansions and a tight linkage with genes encoding a novel and distinct family of predicted transposases, and a member of the Maelstrom-like HMG family. We propose that these fungal members are part of a mobile transposon. To the best of our knowledge, this is the first report of a eukaryotic transposable element that encodes its own DNA-modification enzyme with a potential regulatory role. Through a wider analysis of other poorly characterized DNA-modifying enzymes we also show that the phage Mu Mom-like proteins, which catalyze the N6-carbamoylmethylation of adenines, are also linked to diverse families of bacterial transposases, suggesting that DNA modification by transposable elements might have a more general presence than previously appreciated. Among the other families of 2-oxoglutarate- and iron(II)-dependent dioxygenases identified in this study, one which is found in algae, is predicted to mainly comprise of RNA-modifying enzymes and shows a striking diversity in protein domain architectures suggesting the presence of RNA modifications with possibly unique adaptive roles. The results presented here are likely to provide the means for future investigation of unexpected epigenetic modifications, such as hydroxymethyl cytosine, that could profoundly impact our understanding of gene regulation and processes such as DNA demethylation.
Figures

Similar articles
-
Computational identification of novel biochemical systems involved in oxidation, glycosylation and other complex modifications of bases in DNA.Nucleic Acids Res. 2013 Sep;41(16):7635-55. doi: 10.1093/nar/gkt573. Epub 2013 Jun 28. Nucleic Acids Res. 2013. PMID: 23814188 Free PMC article.
-
Lineage-specific expansions of TET/JBP genes and a new class of DNA transposons shape fungal genomic and epigenetic landscapes.Proc Natl Acad Sci U S A. 2014 Feb 4;111(5):1676-83. doi: 10.1073/pnas.1321818111. Epub 2014 Jan 7. Proc Natl Acad Sci U S A. 2014. PMID: 24398522 Free PMC article.
-
Natural history of eukaryotic DNA methylation systems.Prog Mol Biol Transl Sci. 2011;101:25-104. doi: 10.1016/B978-0-12-387685-0.00002-0. Prog Mol Biol Transl Sci. 2011. PMID: 21507349
-
Introduction: Metals in Biology: α-Ketoglutarate/Iron-Dependent Dioxygenases.J Biol Chem. 2015 Aug 21;290(34):20700-20701. doi: 10.1074/jbc.R115.675652. Epub 2015 Jul 7. J Biol Chem. 2015. PMID: 26152720 Free PMC article. Review.
-
ALKBHs-facilitated RNA modifications and de-modifications.DNA Repair (Amst). 2016 Aug;44:87-91. doi: 10.1016/j.dnarep.2016.05.026. Epub 2016 May 17. DNA Repair (Amst). 2016. PMID: 27237585 Free PMC article. Review.
Cited by
-
Tet family of 5-methylcytosine dioxygenases in mammalian development.J Hum Genet. 2013 Jul;58(7):421-7. doi: 10.1038/jhg.2013.63. Epub 2013 May 30. J Hum Genet. 2013. PMID: 23719188 Free PMC article. Review.
-
Distinct and stage-specific contributions of TET1 and TET2 to stepwise cytosine oxidation in the transition from naive to primed pluripotency.Sci Rep. 2020 Jul 21;10(1):12066. doi: 10.1038/s41598-020-68600-3. Sci Rep. 2020. PMID: 32694513 Free PMC article.
-
Alterations of metabolic genes and metabolites in cancer.Semin Cell Dev Biol. 2012 Jun;23(4):370-80. doi: 10.1016/j.semcdb.2012.01.013. Epub 2012 Jan 28. Semin Cell Dev Biol. 2012. PMID: 22306135 Free PMC article. Review.
-
DNA methylation, its mediators and genome integrity.Int J Biol Sci. 2015 Apr 8;11(5):604-17. doi: 10.7150/ijbs.11218. eCollection 2015. Int J Biol Sci. 2015. PMID: 25892967 Free PMC article. Review.
-
Tet family proteins and 5-hydroxymethylcytosine in development and disease.Development. 2012 Jun;139(11):1895-902. doi: 10.1242/dev.070771. Development. 2012. PMID: 22569552 Free PMC article. Review.
References
-
- Bloomfield VA, Crothers DM, Tinoco I., Jr . Nucleic Acids: Structures, Properties and Functions. Sausalito, CA: University Science Books; 2000.
-
- Pfeifer GP. Mutagenesis at methylated CpG sequences. Curr Top Microbiol Immunol. 2006;301:259–81. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials