

Propriospinal bypass of the serotonergic system that can facilitate stepping
- PMID: 19403834
- PMCID: PMC2940277
- DOI: 10.1523/JNEUROSCI.6058-08.2009
Propriospinal bypass of the serotonergic system that can facilitate stepping
Abstract
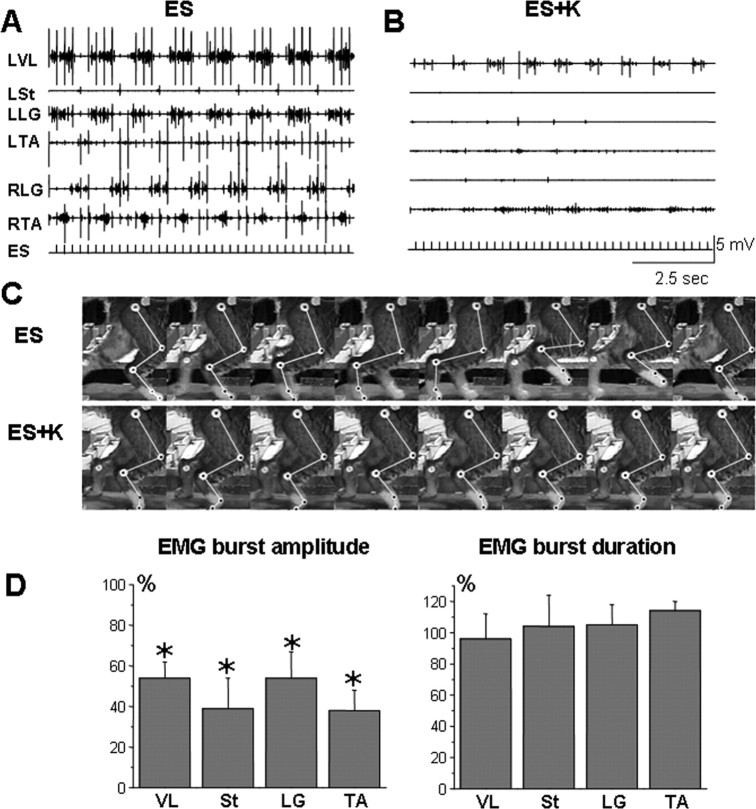
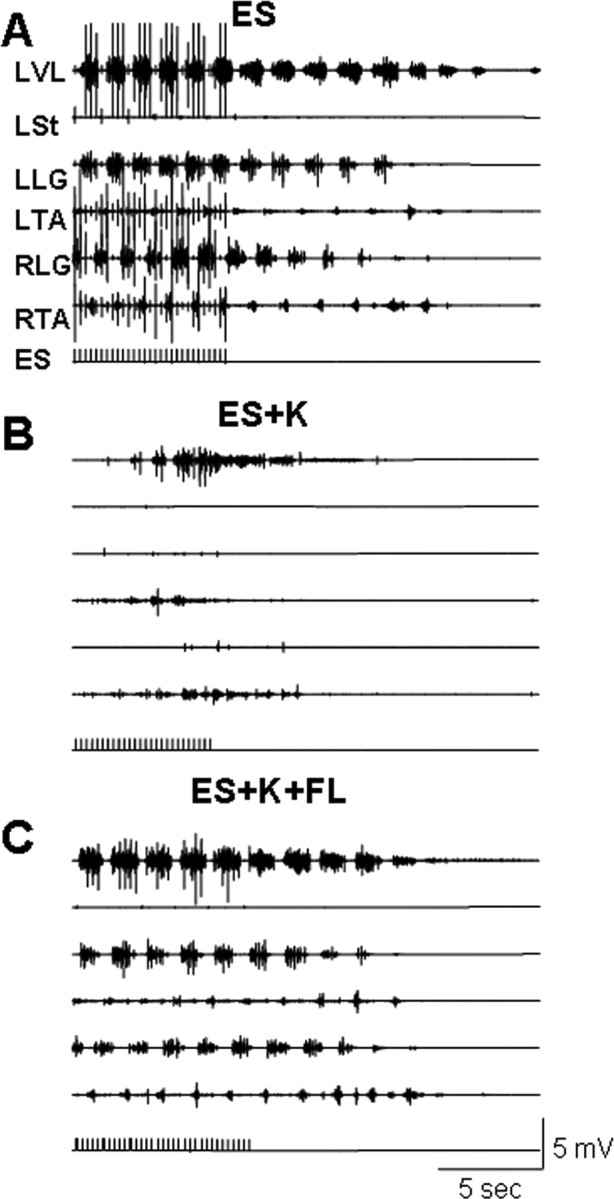
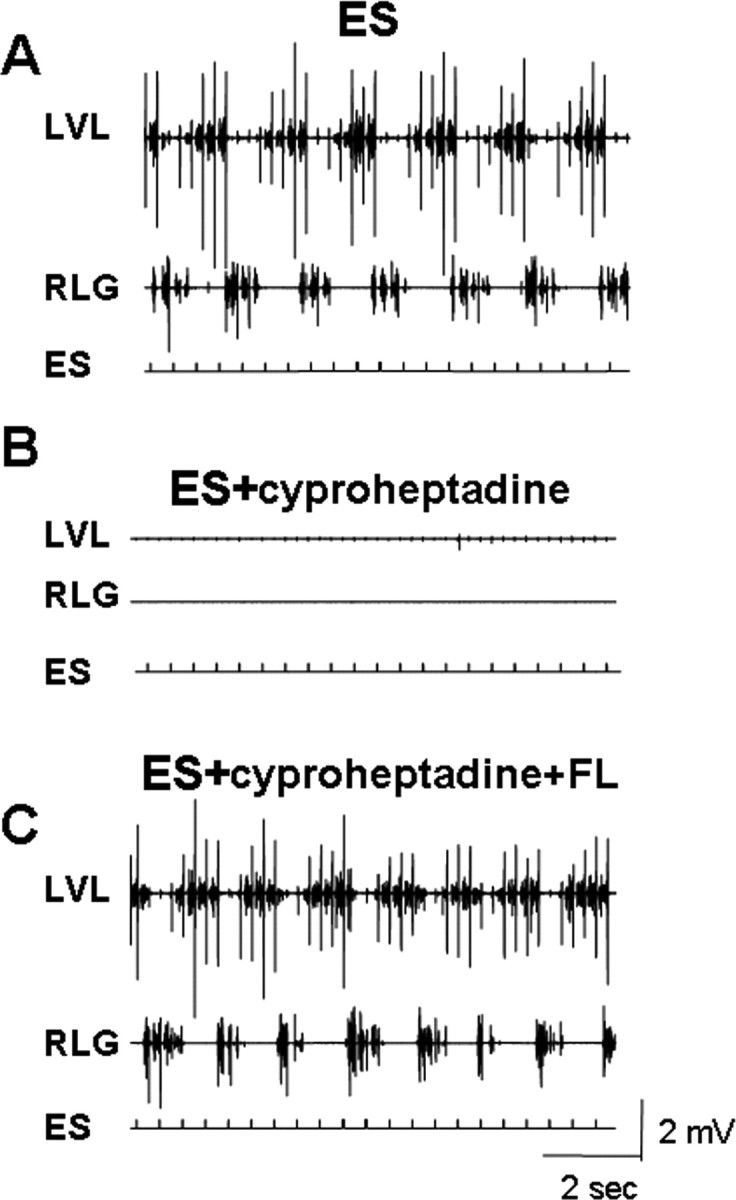
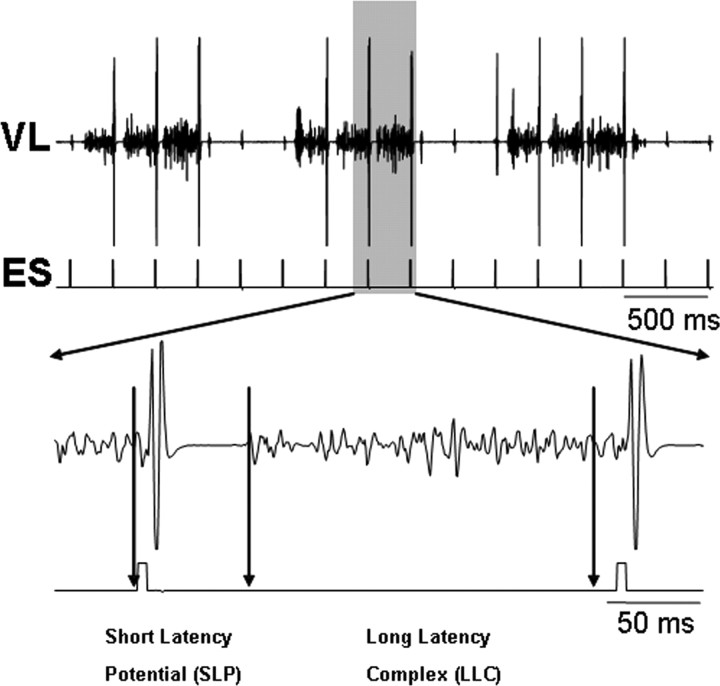
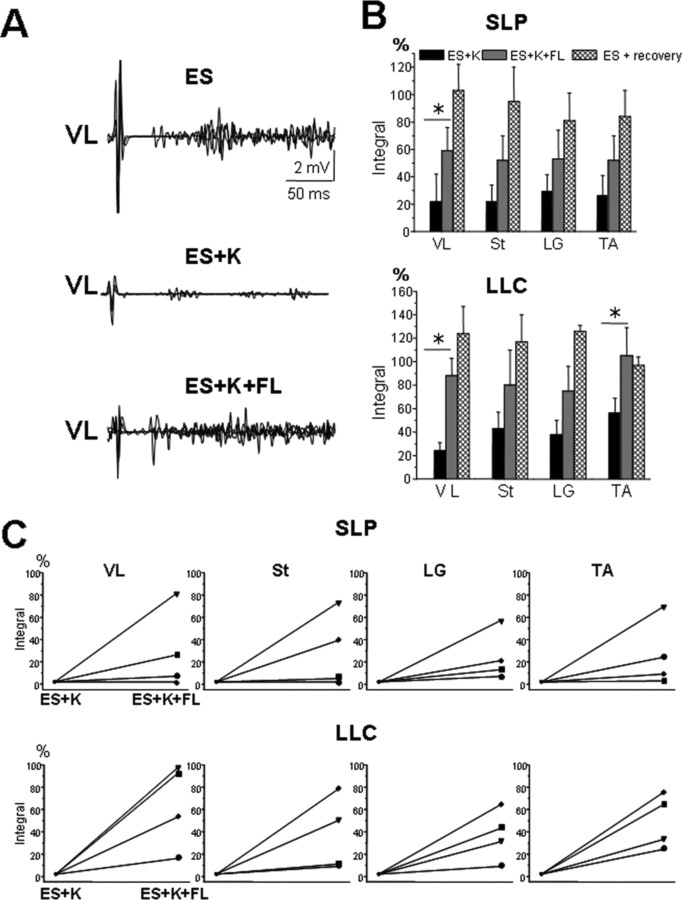
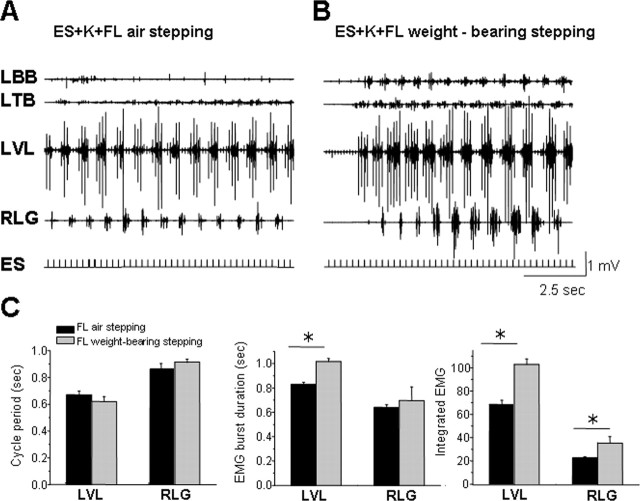
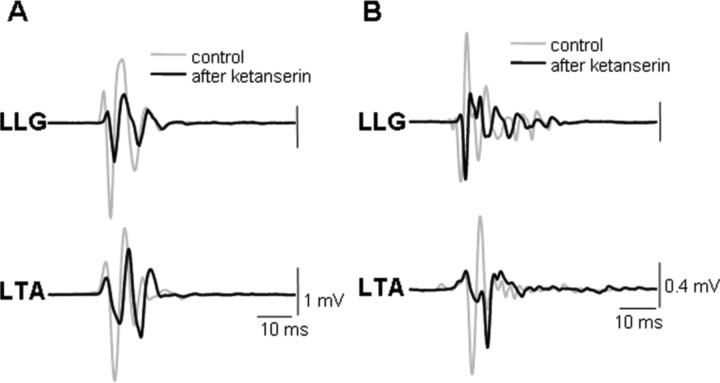
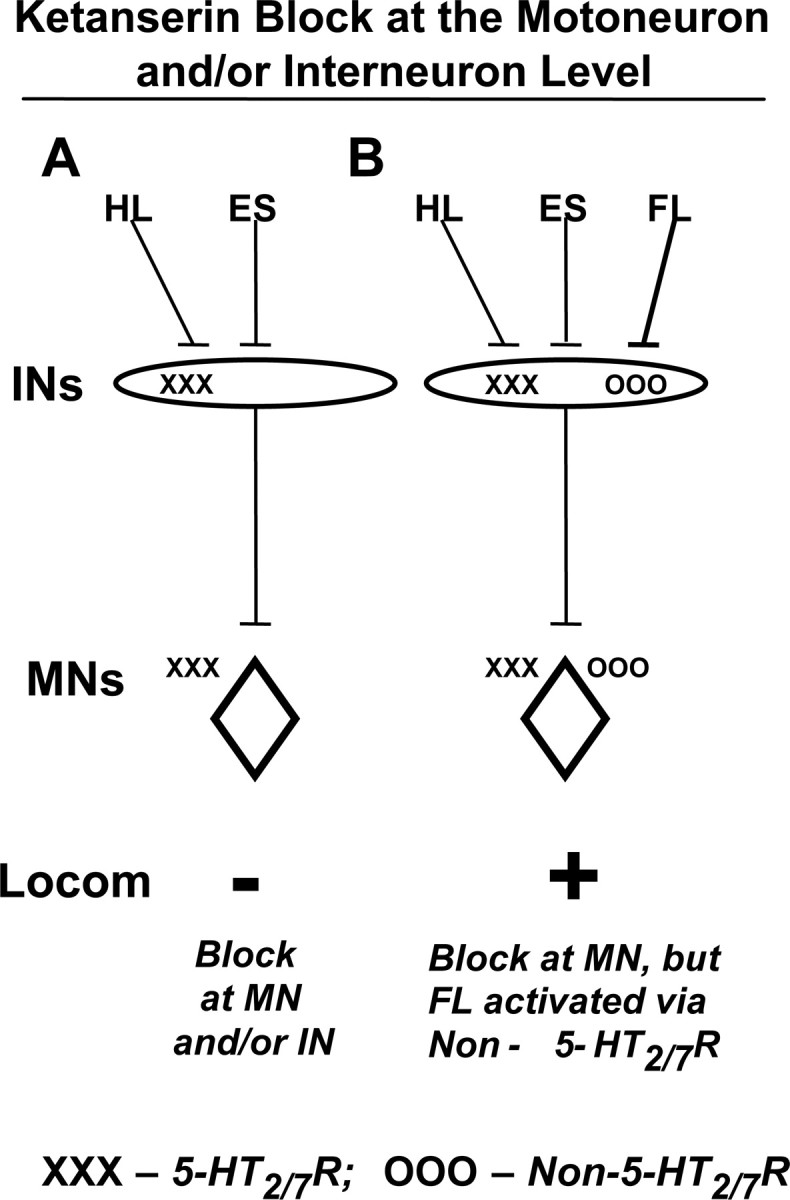
The neurotransmitter systems mediating spinal locomotion in response to epidural spinal cord stimulation (ES) have not been identified. Here, we examine the role of the serotonergic system in regulating locomotor behavior of decerebrated cats during ES at L4-L5. ES elicited coordinated, weight-bearing, hindlimb stepping with plantar foot placement on a moving treadmill belt. Ketanserin [a 5-hydroxytryptamine (serotonin) (5-HT)(2/7) receptor antagonist] depressed this locomotor activity: only weak rhythmic movements without plantar foot placement and depressed EMG activity were observed. Cyproheptadine, a nonselective 5-HT blocker, prevented facilitation of stepping by epidural stimulation. These data demonstrate an important role of the serotonergic system in facilitating locomotion in the presence of epidural stimulation. In the presence of ketanserin, passive movements of one forelimb in a step-like manner immediately induced stepping of both hindlimbs with EMG patterns similar to those observed with ES without ketanserin. Thus, a non-5-HT-dependent spinal circuitry projecting from the cervical to the lumbar region of the spinal cord can facilitate stepping. The specific neurotransmitters responsible for this forelimb-facilitated stepping of the hindlimbs are unknown. These data suggest that a 5-HT(2/7) receptor-dependent pathway that processes hindlimb locomotor-like proprioception to facilitate hindlimb stepping can be complemented with proprioceptive afferents from the forelimbs via a non-5-HT(2/7) receptor neurotransmitter system. Thus, different neurotransmitter receptor systems can be used to mediate the same type of sensory event, i.e., locomotor-like proprioception to facilitate the same motor task, i.e., hindlimb stepping.
Figures
Similar articles
-
Activation of spinal locomotor circuits in the decerebrated cat by spinal epidural and/or intraspinal electrical stimulation.Brain Res. 2015 Mar 10;1600:84-92. doi: 10.1016/j.brainres.2014.11.003. Epub 2014 Nov 12. Brain Res. 2015. PMID: 25446455
-
Facilitation of stepping with epidural stimulation in spinal rats: role of sensory input.J Neurosci. 2008 Jul 30;28(31):7774-80. doi: 10.1523/JNEUROSCI.1069-08.2008. J Neurosci. 2008. PMID: 18667609 Free PMC article.
-
[Significance of peripheral feedback in stepping movement generation under epideral spinal cord stimulation].Ross Fiziol Zh Im I M Sechenova. 2005 Dec;91(12):1407-20. Ross Fiziol Zh Im I M Sechenova. 2005. PMID: 16493922 Russian.
-
Can the mammalian lumbar spinal cord learn a motor task?Med Sci Sports Exerc. 1994 Dec;26(12):1491-7. Med Sci Sports Exerc. 1994. PMID: 7869884 Review.
-
The role of serotonin in the control of locomotor movements and strategies for restoring locomotion after spinal cord injury.Acta Neurobiol Exp (Wars). 2014;74(2):172-87. doi: 10.55782/ane-2014-1983. Acta Neurobiol Exp (Wars). 2014. PMID: 24993627 Review.
Cited by
-
Simultaneous bidirectional hindlimb locomotion in decerebrate cats.Sci Rep. 2021 Feb 5;11(1):3252. doi: 10.1038/s41598-021-82722-2. Sci Rep. 2021. PMID: 33547397 Free PMC article.
-
ROLE OF FORELIMB MORPHOLOGY IN MUSCLE SENSORIMOTOR FUNCTIONS DURING LOCOMOTION IN THE CAT.bioRxiv [Preprint]. 2024 Jul 16:2024.07.11.603106. doi: 10.1101/2024.07.11.603106. bioRxiv. 2024. Update in: J Physiol. 2025 Jan;603(2):447-487. doi: 10.1113/JP287448. PMID: 39071389 Free PMC article. Updated. Preprint.
-
Distribution of Spinal Neuronal Networks Controlling Forward and Backward Locomotion.J Neurosci. 2018 May 16;38(20):4695-4707. doi: 10.1523/JNEUROSCI.2951-17.2018. Epub 2018 Apr 20. J Neurosci. 2018. PMID: 29678875 Free PMC article.
-
Integrating multiple sensory systems to modulate neural networks controlling posture.J Neurophysiol. 2015 Dec;114(6):3306-14. doi: 10.1152/jn.00583.2015. Epub 2015 Oct 7. J Neurophysiol. 2015. PMID: 26445868 Free PMC article.
-
Planar Covariation of Hindlimb and Forelimb Elevation Angles during Terrestrial and Aquatic Locomotion of Dogs.PLoS One. 2015 Jul 28;10(7):e0133936. doi: 10.1371/journal.pone.0133936. eCollection 2015. PLoS One. 2015. PMID: 26218076 Free PMC article.
References
-
- Akay T, McVea DA, Tachibana A, Pearson KG. Coordination of fore and hind leg stepping in cats on a transversely-split treadmill. Exp Brain Res. 2006;175:211–222. - PubMed
-
- Antri M, Orsal D, Barthe JY. Locomotor recovery in the chronic spinal rat: effects of long-term treatment with a 5-HT2 agonist. Eur J Neurosci. 2002;16:467–476. - PubMed
-
- Ballion B, Morin D, Viala D. Forelimb locomotor generators and quadrupedal locomotion in the neonatal rat. Eur J Neurosci. 2001;14:1727–1738. - PubMed
-
- Barbeau H, Rossignol S. The effects of serotonergic drugs on the locomotor pattern and on cutaneous reflexes of the adult chronic spinal cat. Brain Res. 1990;514:55–67. - PubMed
-
- Barbeau H, Rossignol S. Initiation and modulation of locomotor pattern in the adult chronic spinal cat by noradrenergic, serotonergic and dopaminergic drugs. Brain Res. 1991;546:250–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
