

K63-linked ubiquitin chains as a specific signal for protein sorting into the multivesicular body pathway
- PMID: 19398763
- PMCID: PMC2700384
- DOI: 10.1083/jcb.200810114
K63-linked ubiquitin chains as a specific signal for protein sorting into the multivesicular body pathway
Abstract
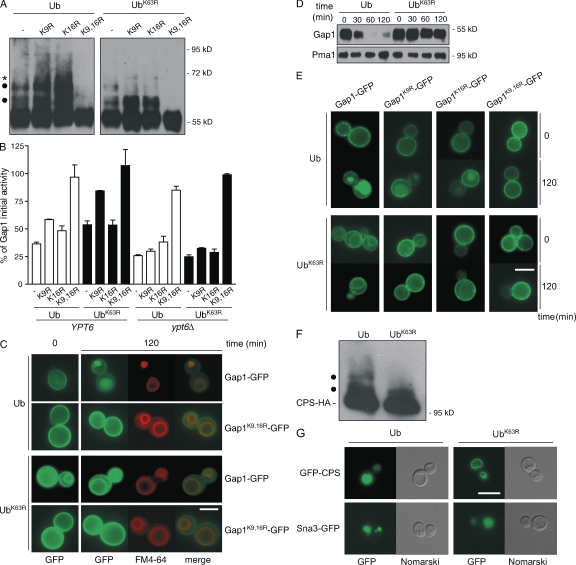
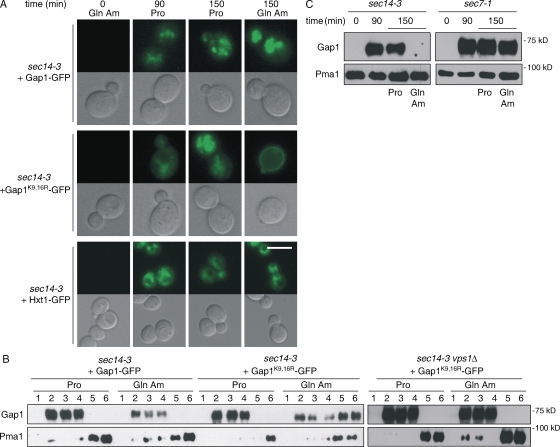
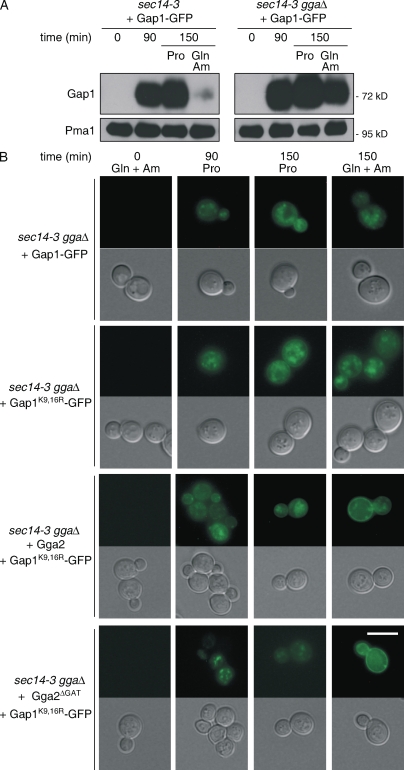
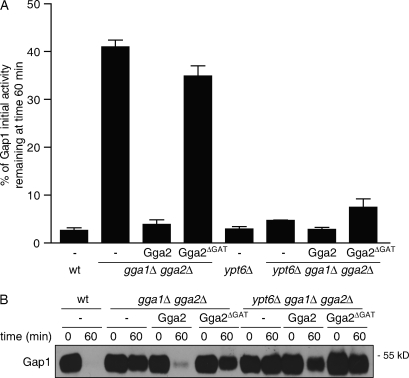
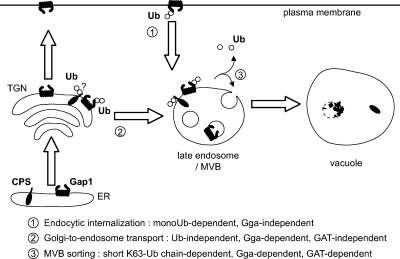
A growing number of yeast and mammalian plasma membrane proteins are reported to be modified with K63-linked ubiquitin (Ub) chains. However, the relative importance of this modification versus monoubiquitylation in endocytosis, Golgi to endosome traffic, and sorting into the multivesicular body (MVB) pathway remains unclear. In this study, we show that K63-linked ubiquitylation of the Gap1 permease is essential for its entry into the MVB pathway. Carboxypeptidase S also requires modification with a K63-Ub chain for correct MVB sorting. In contrast, monoubiquitylation of a single target lysine of Gap1 is a sufficient signal for its internalization from the cell surface, and Golgi to endosome transport of the permease requires neither its ubiquitylation nor the Ub-binding GAT (Gga and Tom1) domain of Gga (Golgi localizing, gamma-ear containing, ARF binding) adapter proteins, the latter being crucial for subsequent MVB sorting of the permease. Our data reveal that K63-linked Ub chains act as a specific signal for MVB sorting, providing further insight into the Ub code of membrane protein trafficking.
Figures
Similar articles
-
The GAT domains of clathrin-associated GGA proteins have two ubiquitin binding motifs.J Biol Chem. 2004 Dec 24;279(52):54808-16. doi: 10.1074/jbc.M406654200. Epub 2004 Oct 19. J Biol Chem. 2004. PMID: 15494413 Free PMC article.
-
Evidence for a direct role of the Doa4 deubiquitinating enzyme in protein sorting into the MVB pathway.Traffic. 2007 May;8(5):566-81. doi: 10.1111/j.1600-0854.2007.00553.x. Epub 2007 Mar 21. Traffic. 2007. PMID: 17376168
-
Ubiquitin is required for sorting to the vacuole of the yeast general amino acid permease, Gap1.J Biol Chem. 2001 Nov 23;276(47):43949-57. doi: 10.1074/jbc.M102945200. Epub 2001 Aug 10. J Biol Chem. 2001. PMID: 11500494
-
The ubiquitin code of yeast permease trafficking.Trends Cell Biol. 2010 Apr;20(4):196-204. doi: 10.1016/j.tcb.2010.01.004. Trends Cell Biol. 2010. PMID: 20138522 Review.
-
Versatile role of the yeast ubiquitin ligase Rsp5p in intracellular trafficking.Biochem Soc Trans. 2008 Oct;36(Pt 5):791-6. doi: 10.1042/BST0360791. Biochem Soc Trans. 2008. PMID: 18793138 Review.
Cited by
-
A calcineurin-dependent switch controls the trafficking function of α-arrestin Aly1/Art6.J Biol Chem. 2013 Aug 16;288(33):24063-80. doi: 10.1074/jbc.M113.478511. Epub 2013 Jul 3. J Biol Chem. 2013. PMID: 23824189 Free PMC article.
-
Ataxin-3 regulates aggresome formation of copper-zinc superoxide dismutase (SOD1) by editing K63-linked polyubiquitin chains.J Biol Chem. 2012 Aug 17;287(34):28576-85. doi: 10.1074/jbc.M111.299990. Epub 2012 Jul 3. J Biol Chem. 2012. PMID: 22761419 Free PMC article.
-
Bimodal antagonism of PKA signalling by ARHGAP36.Nat Commun. 2016 Oct 7;7:12963. doi: 10.1038/ncomms12963. Nat Commun. 2016. PMID: 27713425 Free PMC article.
-
The de-ubiquitylating enzyme DUBA is essential for spermatogenesis in Drosophila.Cell Death Differ. 2016 Dec;23(12):2019-2030. doi: 10.1038/cdd.2016.79. Epub 2016 Aug 12. Cell Death Differ. 2016. PMID: 27518434 Free PMC article.
-
Targeting the Lysosomal Degradation of Rab22a-NeoF1 Fusion Protein for Osteosarcoma Lung Metastasis.Adv Sci (Weinh). 2023 Feb;10(5):e2205483. doi: 10.1002/advs.202205483. Epub 2022 Dec 18. Adv Sci (Weinh). 2023. PMID: 36529692 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
