

Antigen sensitivity is a major determinant of CD8+ T-cell polyfunctionality and HIV-suppressive activity
- PMID: 19389882
- PMCID: PMC2710928
- DOI: 10.1182/blood-2009-02-206557
Antigen sensitivity is a major determinant of CD8+ T-cell polyfunctionality and HIV-suppressive activity
Abstract
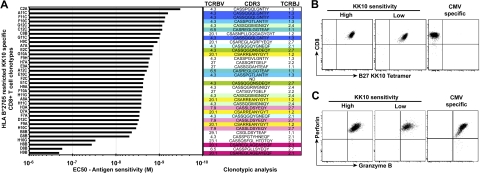
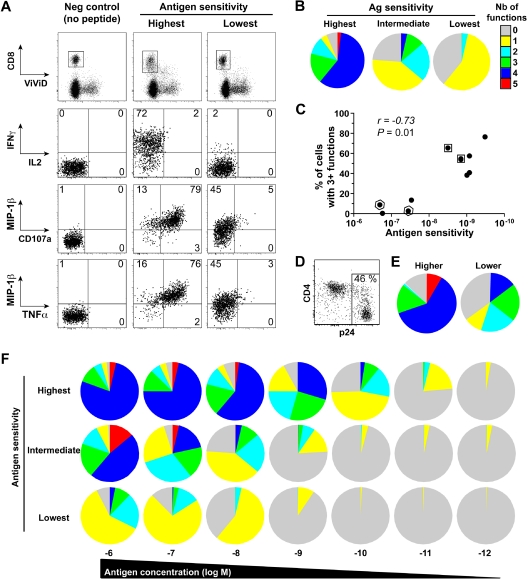
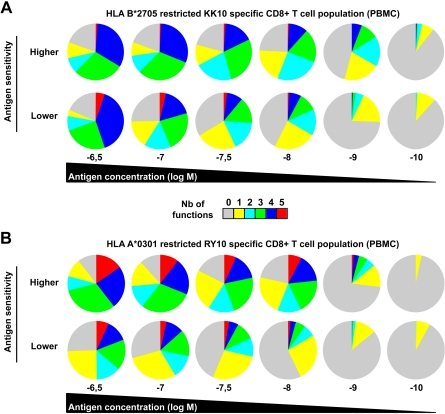
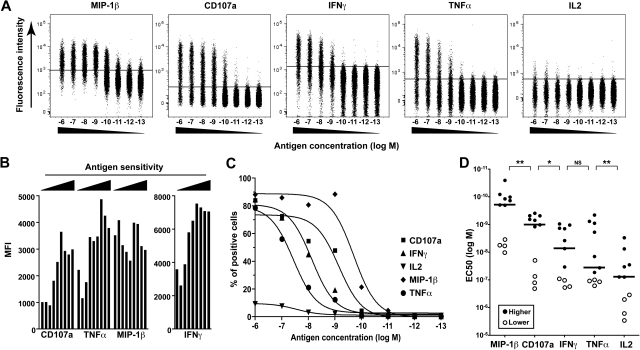
CD8(+) T cells are major players in the immune response against HIV. However, recent failures in the development of T cell-based vaccines against HIV-1 have emphasized the need to reassess our basic knowledge of T cell-mediated efficacy. CD8(+) T cells from HIV-1-infected patients with slow disease progression exhibit potent polyfunctionality and HIV-suppressive activity, yet the factors that unify these properties are incompletely understood. We performed a detailed study of the interplay between T-cell functional attributes using a bank of HIV-specific CD8(+) T-cell clones isolated in vitro; this approach enabled us to overcome inherent difficulties related to the in vivo heterogeneity of T-cell populations and address the underlying determinants that synthesize the qualities required for antiviral efficacy. Conclusions were supported by ex vivo analysis of HIV-specific CD8(+) T cells from infected donors. We report that attributes of CD8(+) T-cell efficacy against HIV are linked at the level of antigen sensitivity. Highly sensitive CD8(+) T cells display polyfunctional profiles and potent HIV-suppressive activity. These data provide new insights into the mechanisms underlying CD8(+) T-cell efficacy against HIV, and indicate that vaccine strategies should focus on the induction of HIV-specific T cells with high levels of antigen sensitivity to elicit potent antiviral efficacy.
Figures
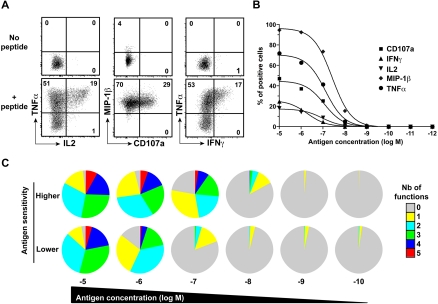
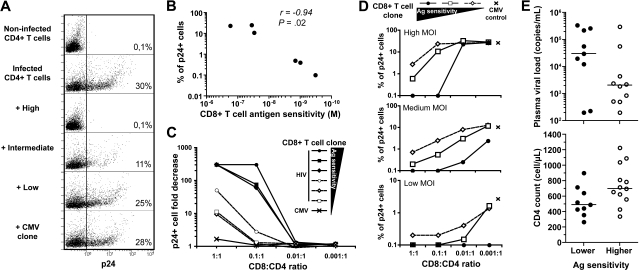
Similar articles
-
Suppression of HIV replication in the resting CD4+ T cell reservoir by autologous CD8+ T cells: implications for the development of therapeutic strategies.Proc Natl Acad Sci U S A. 2001 Jan 2;98(1):253-8. doi: 10.1073/pnas.98.1.253. Proc Natl Acad Sci U S A. 2001. PMID: 11136258 Free PMC article.
-
The link between CD8⁺ T-cell antigen-sensitivity and HIV-suppressive capacity depends on HLA restriction, target epitope and viral isolate.AIDS. 2014 Feb 20;28(4):477-86. doi: 10.1097/QAD.0000000000000175. AIDS. 2014. PMID: 24384691
-
Polyfunctional CD8+ T-Cell Response to Autologous Peptides from Protease and Reverse Transcriptase of HIV-1 Clade B.Curr HIV Res. 2019;17(5):350-359. doi: 10.2174/1570162X17666191017105910. Curr HIV Res. 2019. PMID: 31622220
-
Multiple Approaches for Increasing the Immunogenicity of an Epitope-Based Anti-HIV Vaccine.AIDS Res Hum Retroviruses. 2015 Nov;31(11):1077-88. doi: 10.1089/AID.2015.0101. Epub 2015 Aug 13. AIDS Res Hum Retroviruses. 2015. PMID: 26149745 Review.
-
CD8 lymphocytes in HIV infection: helpful and harmful.J Clin Lab Immunol. 1997;49(1):15-32. J Clin Lab Immunol. 1997. PMID: 9819670 Review.
Cited by
-
Deconvolving heterogeneity in the CD8+ T-cell response to HIV.Curr Opin HIV AIDS. 2012 Jan;7(1):38-43. doi: 10.1097/COH.0b013e32834dde1c. Curr Opin HIV AIDS. 2012. PMID: 22156844 Free PMC article. Review.
-
Public T cell receptors confer high-avidity CD4 responses to HIV controllers.J Clin Invest. 2016 Jun 1;126(6):2093-108. doi: 10.1172/JCI83792. Epub 2016 Apr 25. J Clin Invest. 2016. PMID: 27111229 Free PMC article.
-
Phenotypic and functional profile of HIV-inhibitory CD8 T cells elicited by natural infection and heterologous prime/boost vaccination.J Virol. 2010 May;84(10):4998-5006. doi: 10.1128/JVI.00138-10. Epub 2010 Mar 3. J Virol. 2010. PMID: 20200250 Free PMC article.
-
Considering Genomic and Immunological Correlates of Protection for a Dengue Intervention.Vaccines (Basel). 2019 Dec 3;7(4):203. doi: 10.3390/vaccines7040203. Vaccines (Basel). 2019. PMID: 31816907 Free PMC article. Review.
-
Dendritic cells exposed to MVA-based HIV-1 vaccine induce highly functional HIV-1-specific CD8(+) T cell responses in HIV-1-infected individuals.PLoS One. 2011;6(5):e19644. doi: 10.1371/journal.pone.0019644. Epub 2011 May 18. PLoS One. 2011. PMID: 21625608 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
