

Review
doi: 10.1093/nar/gkp250.
Epub 2009 Apr 21.
Theoretical models of DNA topology simplification by type IIA DNA topoisomerases
Affiliations
- PMID: 19383879
- PMCID: PMC2691845
- DOI: 10.1093/nar/gkp250
Item in Clipboard
Review
Theoretical models of DNA topology simplification by type IIA DNA topoisomerases
Nucleic Acids Res.
2009 Jun.
Abstract
It was discovered 12 years ago that type IIA topoisomerases can simplify DNA topology--the steady-state fractions of knots and links created by the enzymes are many times lower than the corresponding equilibrium fractions. Though this property of the enzymes made clear biological sense, it was not clear how small enzymes could selectively change the topology of very large DNA molecules, since topology is a global property and cannot be determined by a local DNA-protein interaction. A few models, suggested to explain the phenomenon, are analyzed in this review. We also consider experimental data that both support and contravene these models.
Figures
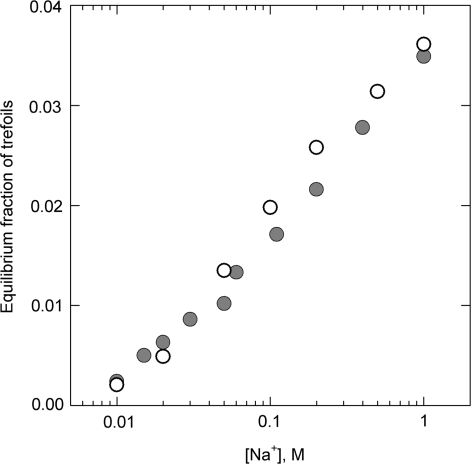
Measured and simulated equilibrium fractions of trefoil knots for different concentrations of sodium ions [based on the data from ref. (9)]. The experiments were performed with 10-kb DNA which was cyclized in solution of different NaCl concentrations via joining the cohesive ends. Each point on the graph (gray circles) is the average of 6–20 determinations. The results of computer simulation, shown by open circles, account for the salt concentration over the effective diameter of the double helix which strongly depends on NaCl concentration (11). Only traces of more complex knots were observed in the experiment.
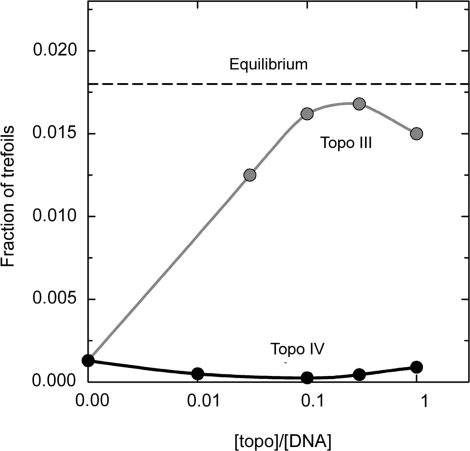
Type IIA topoisomerase, topo IV from E. coli, removes topological links from DNA to level below equilibrium (3). The reaction reached its steady state at substoichiometric values of the enzyme/DNA ratio. The equilibrium value of the knot fraction for the given conditions and DNA length, 7 kb, is shown as a reference level by the dashed line. Also shown, as a control, the fraction of knots found for topo III, type I topoisomerase from E. coli, which does not consume the energy during the catalysis and thus must shift the fraction of trefoils to the equilibrium level.
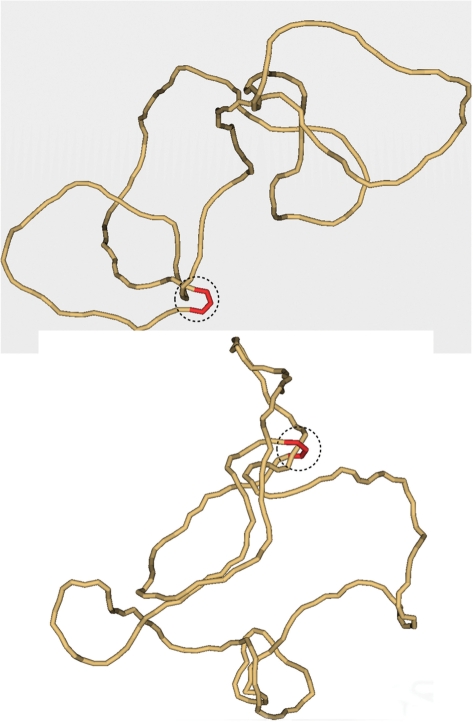
Typical simulated conformation of knotted (top) and unknotted (bottom) 7-kb DNA molecules. Each of the shown conformations has a segment located inside the hairpin-like G segment (red). For both conformations the potential T segment and G segment, which could interact with the enzyme, are circled by the dashed line. It seems clear from the figure that the mutual path of the segments inside the circle cannot specify topology of the entire chains. Indeed, the topology of both conformations can be easily changed outside the dashed circles. The conformations were selected from the equilibrium ensemble generated by a Metropolis Monte Carlo procedure (5).
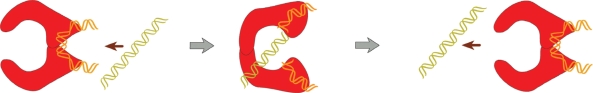
The model of type IIA topoisomerase action. The enzyme (red) bends the G segment of DNA into a hairpin-like conformation. The entrance gate for the T segment of DNA is inside the hairpin. Thus, the T segment can pass through the G segment only from inside to outside the hairpin. Although it is not clear that the suggested mechanism has to provide simplification of DNA topology, the computational analysis shows that it really does (5).
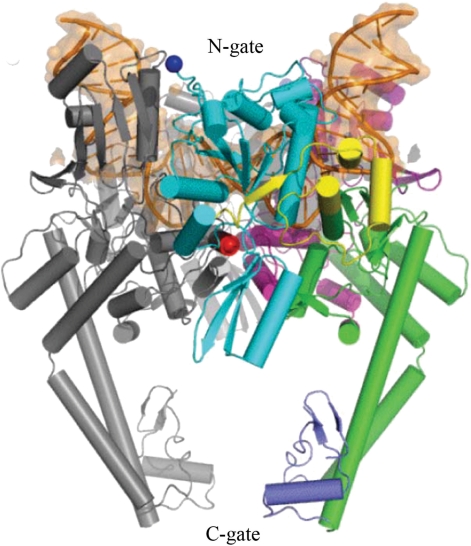
Cartoon representation of the structure of the Topo II DNA-binding and cleavage core bound with DNA fragment [reproduced from ref. (18)]. DNA fragment, shown by orange, is bent by 150° in the complex.
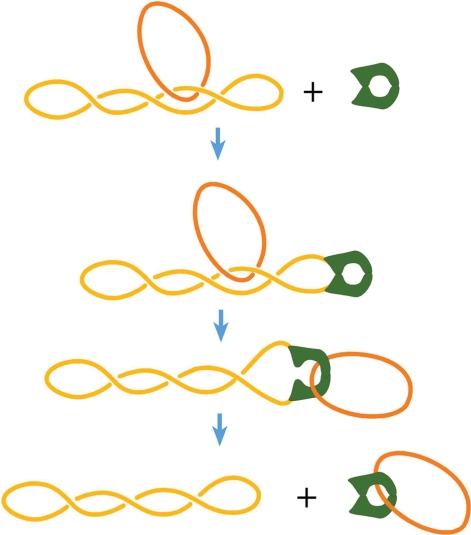
The two-gate mechanism of the strand-passing reaction. The diagram illustrates the experiment with mutant yeast Topo II enzyme, which the C-gate was locked by disulfide links (13). The reaction substrate consisted of the large supercoiled molecule linked with small nicked circular DNA. Unlinking of circular DNA molecules accompanied by the AMPPNP driven closure of the N-gate, left the small circular molecule topologically linked with mutant Topo II. Such an outcome is possible only if a T segment enters through the N-gate of the enzyme and exits through the C-gate.

Diagram of a hooked juxtaposition of two DNA segments. It was suggested that such juxtapositions are typical among segment juxtaposition in linked and knotted molecules and represent a binding substrate for type IIA topoisomerases (6).
Similar articles
-
How do type II topoisomerases use ATP hydrolysis to simplify DNA topology beyond equilibrium? Investigating the relaxation reaction of nonsupercoiling type II topoisomerases.J Mol Biol. 2009 Feb 6;385(5):1397-408. doi: 10.1016/j.jmb.2008.11.056. Epub 2008 Dec 7. J Mol Biol. 2009. PMID: 19094994 Free PMC article.
-
Simplification of DNA topology below equilibrium values by type II topoisomerases.Science. 1997 Aug 1;277(5326):690-3. doi: 10.1126/science.277.5326.690. Science. 1997. PMID: 9235892
-
Local sensing of global DNA topology: from crossover geometry to type II topoisomerase processivity.Nucleic Acids Res. 2011 Nov 1;39(20):8665-76. doi: 10.1093/nar/gkr556. Epub 2011 Jul 15. Nucleic Acids Res. 2011. PMID: 21764774 Free PMC article.
-
The role of ATP in the reactions of type II DNA topoisomerases.Biochem Soc Trans. 2010 Apr;38(2):438-42. doi: 10.1042/BST0380438. Biochem Soc Trans. 2010. PMID: 20298198 Review.
-
Simulation of DNA catenanes.Phys Chem Chem Phys. 2009 Dec 7;11(45):10543-52. doi: 10.1039/b910812b. Epub 2009 Oct 23. Phys Chem Chem Phys. 2009. PMID: 20145800 Free PMC article. Review.
Cited by
-
Kinetic pathways of topology simplification by Type-II topoisomerases in knotted supercoiled DNA.Nucleic Acids Res. 2019 Jan 10;47(1):69-84. doi: 10.1093/nar/gky1174. Nucleic Acids Res. 2019. PMID: 30476194 Free PMC article.
-
The Dynamic Interplay Between DNA Topoisomerases and DNA Topology.Biophys Rev. 2016 Sep;8(3):221-231. doi: 10.1007/s12551-016-0206-x. Epub 2016 Jul 2. Biophys Rev. 2016. PMID: 27942270 Free PMC article.
-
The fractal globule as a model of chromatin architecture in the cell.Chromosome Res. 2011 Jan;19(1):37-51. doi: 10.1007/s10577-010-9177-0. Chromosome Res. 2011. PMID: 21274616 Free PMC article.
-
The dynamic interplay between DNA topoisomerases and DNA topology.Biophys Rev. 2016 Nov;8(Suppl 1):101-111. doi: 10.1007/s12551-016-0240-8. Epub 2016 Nov 14. Biophys Rev. 2016. PMID: 28510219 Free PMC article. Review.
-
Promoter polymorphisms of TOP2A and ERCC1 genes as predictive factors for chemotherapy in non-small cell lung cancer patients.Cancer Med. 2020 Jan;9(2):605-614. doi: 10.1002/cam4.2743. Epub 2019 Dec 3. Cancer Med. 2020. PMID: 31797573 Free PMC article.
References
-
- Wang JC. Moving one DNA double helix through another by a type II DNA topoisomerase: the story of a simple molecular machine. Q. Rev. Biophys. 1998;31:107–144. - PubMed
-
- Schoeffler AJ, Berger JM. Recent advances in understanding structure-function relationships in the type II topoisomerase mechanism. Biochem. Soc. Trans. 2005;33:1465–1470. - PubMed
-
- Rybenkov VV, Ullsperger C, Vologodskii AV, Cozzarelli NR. Simplification of DNA topology below equilibrium values by type II topoisomerases. Science. 1997;277:690–693. - PubMed
-
- Yan J, Magnasco MO, Marko JF. A kinetic proofreading mechanism for disentanglement of DNA by topoisomerases. Nature. 1999;401:932–935. - PubMed
