

Analyses of regenerative wave patterns in adult hair follicle populations reveal macro-environmental regulation of stem cell activity
- PMID: 19378257
- PMCID: PMC2759942
- DOI: 10.1387/ijdb.072564mp
Analyses of regenerative wave patterns in adult hair follicle populations reveal macro-environmental regulation of stem cell activity
Abstract
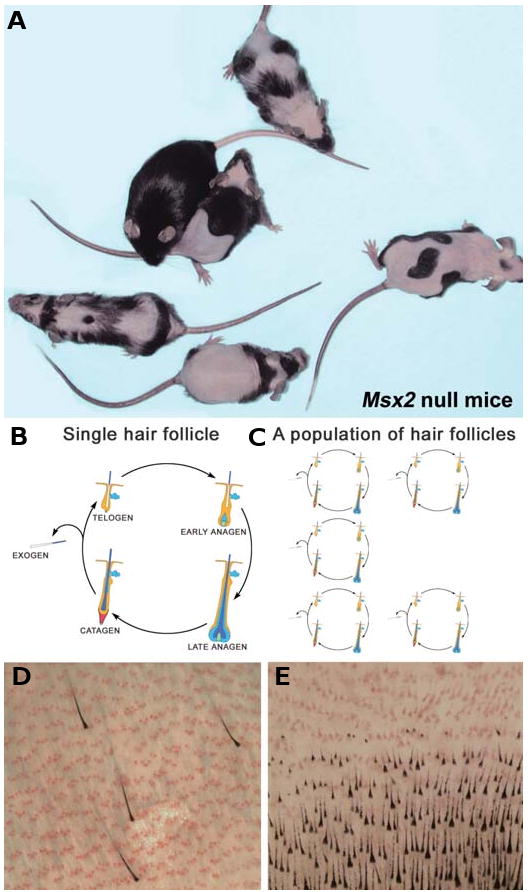
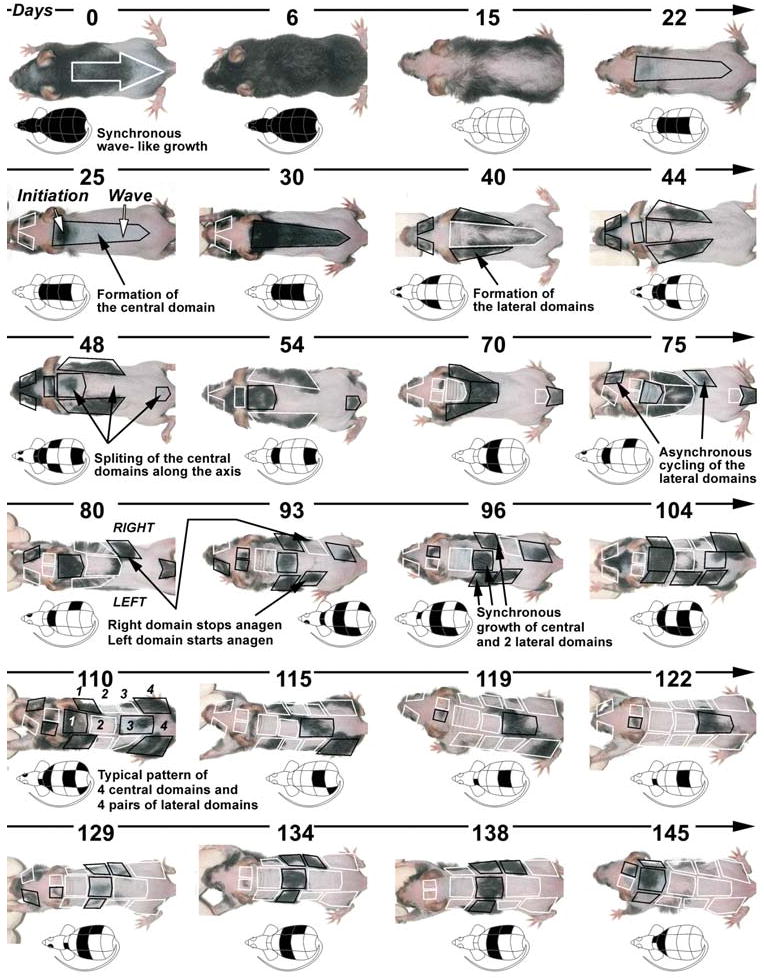
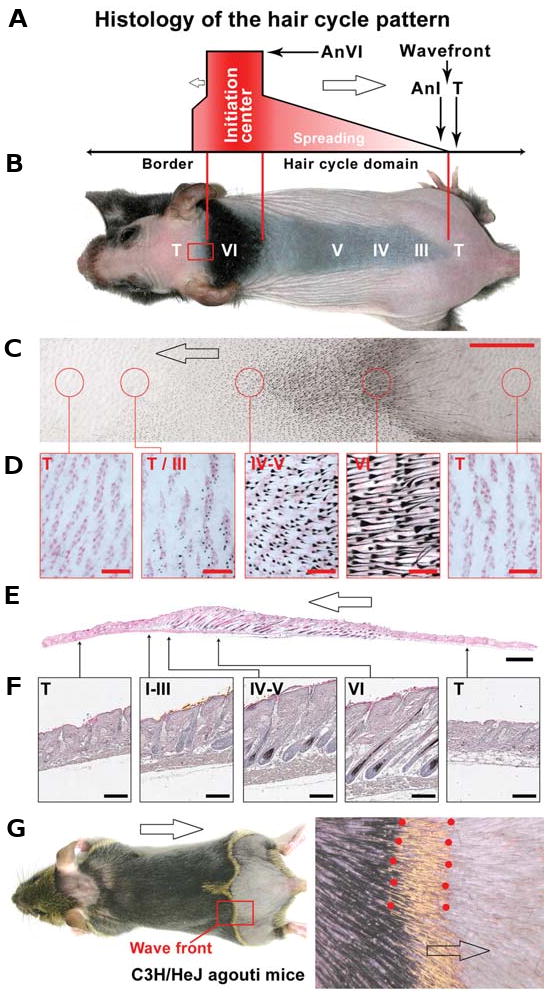
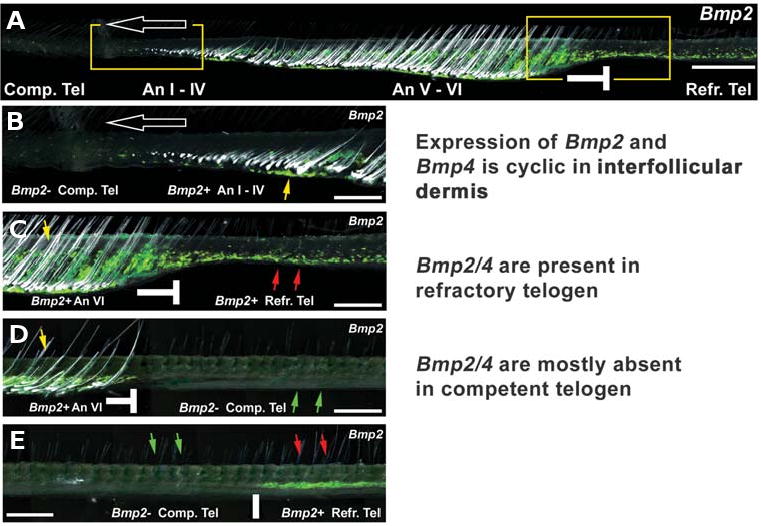
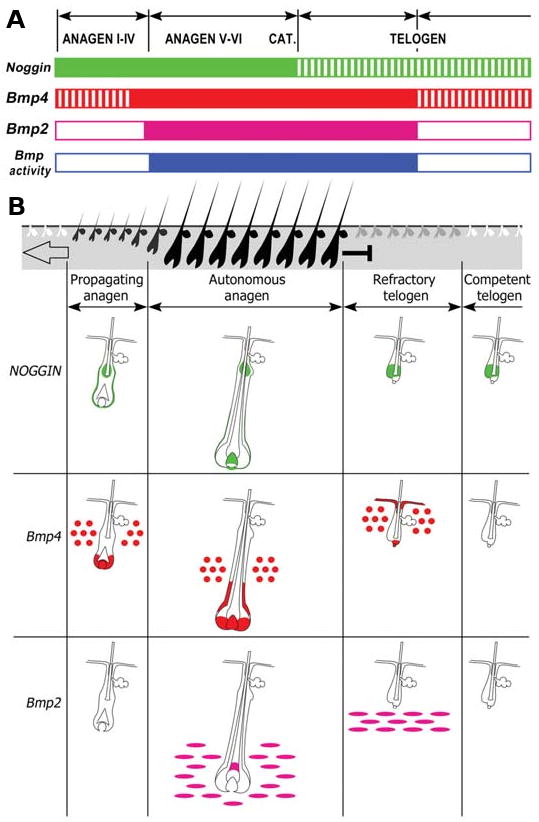
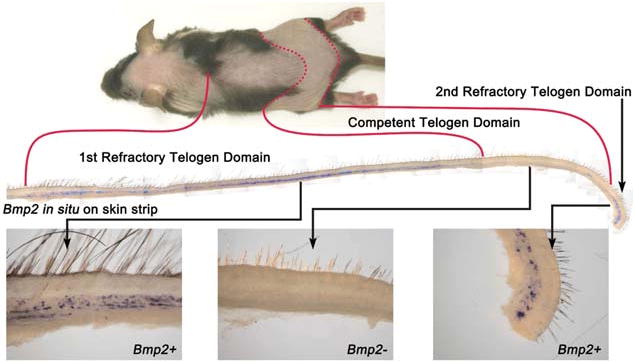
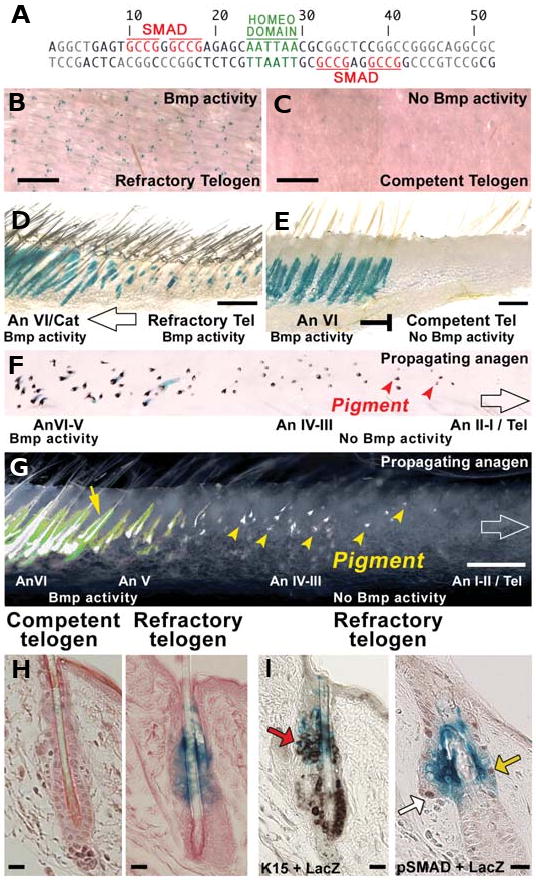
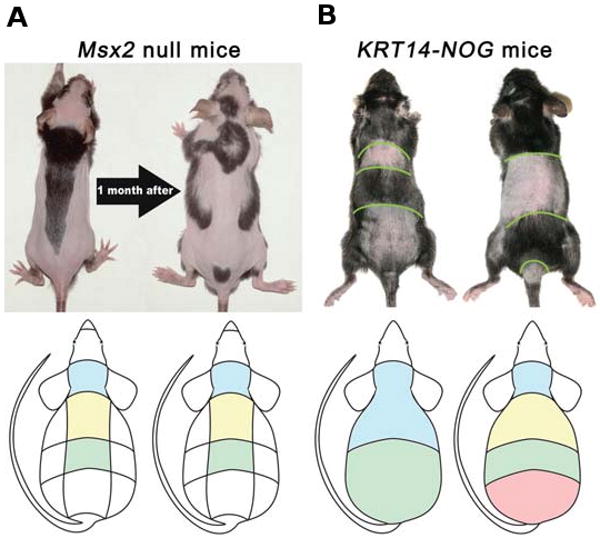
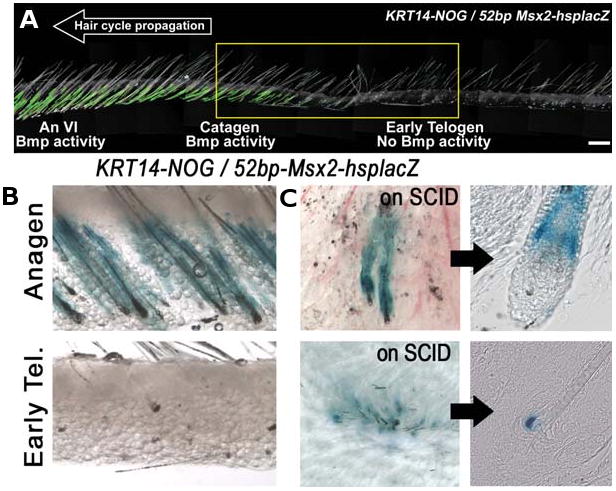
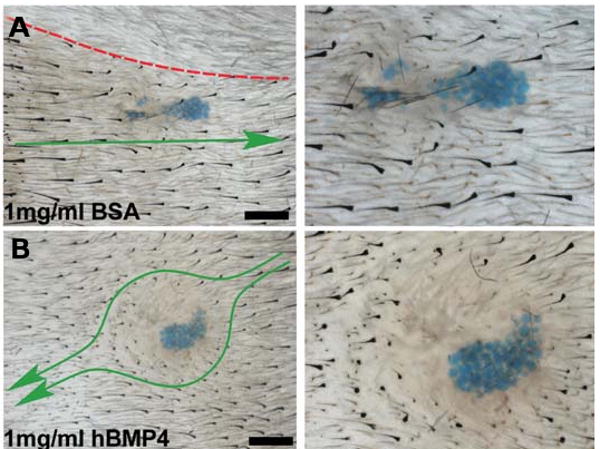
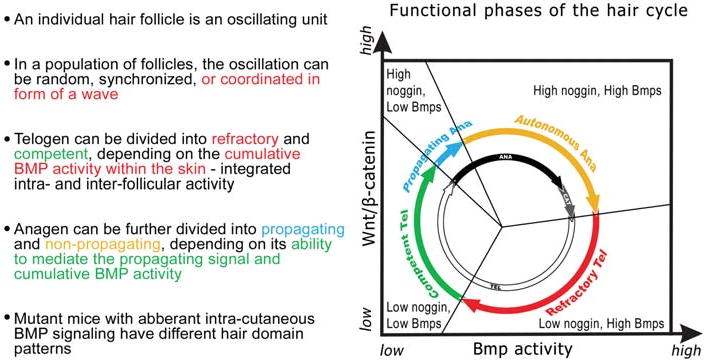
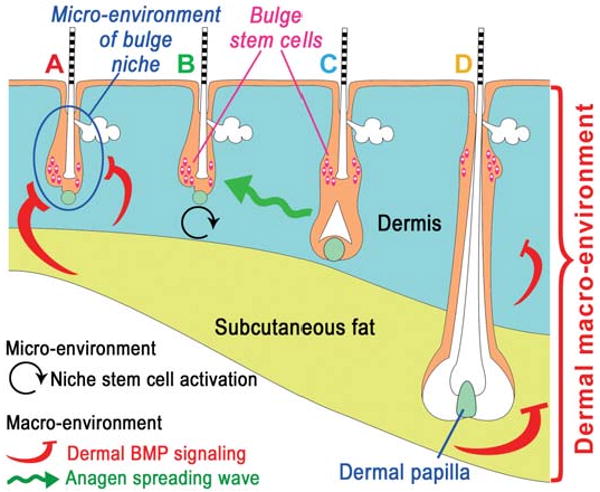
The control of hair growth in the adult mammalian coat is a fascinating topic which has just begun to be explored with molecular genetic tools. Complex hair cycle domains and regenerative hair waves are present in normal adult (> 2 month) mice, but more apparent in mutants with cyclic alopecia phenotypes. Each hair cycle domain consists of initiation site(s), a propagating wave and boundaries. By analyzing the dynamics of hair growth, time required for regeneration after plucking, in situ hybridization and reporter activity, we showed that there is oscillation of intra-follicular Wnt signaling which is synchronous with hair cycling, and there is oscillation of dermal bone morphogenetic protein (BMP) signaling which is asynchronous with hair cycling. The interactions of these two rhythms lead to the recognition of refractory and competent phases in the telogen, and autonomous and propagating phases in the anagen. Boundaries form when propagating anagen waves reach follicles which are in refractory telogen. Experiments showed that Krt14-Nog mice have shortened refractory telogen and simplified wave dynamics. Krt14-Nog skin grafts exhibit non-autonomous interactions with surrounding host skin. Implantation of BMP coated beads into competent telogen skin prevents hair wave propagation around the bead. Thus, we have developed a new molecular understanding of the classic early concepts of inhibitory "chalone", suggesting that stem cells within the hair follicle micro-environment, or other organs, are subject to a higher level of macro-environmental regulation. Such a novel understanding has important implications in the field of regenerative medicine. The unexpected links with Bmp2 expression in subcutaneous adipocytes has implications for systems biology and Evo-Devo.
Figures
Comment in
-
Pattern formation today.Int J Dev Biol. 2009;53(5-6):653-8. doi: 10.1387/ijdb.082594cc. Int J Dev Biol. 2009. PMID: 19557673 Free PMC article. Review.
Similar articles
-
Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration.Nature. 2008 Jan 17;451(7176):340-4. doi: 10.1038/nature06457. Nature. 2008. PMID: 18202659 Free PMC article.
-
Complex hair cycle domain patterns and regenerative hair waves in living rodents.J Invest Dermatol. 2008 May;128(5):1071-80. doi: 10.1038/sj.jid.5701180. J Invest Dermatol. 2008. PMID: 18094733 Free PMC article. Review.
-
Resting no more: re-defining telogen, the maintenance stage of the hair growth cycle.Biol Rev Camb Philos Soc. 2015 Nov;90(4):1179-96. doi: 10.1111/brv.12151. Epub 2014 Nov 19. Biol Rev Camb Philos Soc. 2015. PMID: 25410793 Free PMC article. Review.
-
Regenerative hair waves in aging mice and extra-follicular modulators follistatin, dkk1, and sfrp4.J Invest Dermatol. 2014 Aug;134(8):2086-2096. doi: 10.1038/jid.2014.139. Epub 2014 Mar 11. J Invest Dermatol. 2014. PMID: 24618599 Free PMC article.
-
mTOR signaling promotes stem cell activation via counterbalancing BMP-mediated suppression during hair regeneration.J Mol Cell Biol. 2015 Feb;7(1):62-72. doi: 10.1093/jmcb/mjv005. Epub 2015 Jan 20. J Mol Cell Biol. 2015. PMID: 25609845
Cited by
-
Therapeutic strategy for hair regeneration: hair cycle activation, niche environment modulation, wound-induced follicle neogenesis, and stem cell engineering.Expert Opin Biol Ther. 2013 Mar;13(3):377-91. doi: 10.1517/14712598.2013.739601. Epub 2013 Jan 5. Expert Opin Biol Ther. 2013. PMID: 23289545 Free PMC article. Review.
-
Modelling hair follicle growth dynamics as an excitable medium.PLoS Comput Biol. 2012;8(12):e1002804. doi: 10.1371/journal.pcbi.1002804. Epub 2012 Dec 20. PLoS Comput Biol. 2012. PMID: 23284275 Free PMC article.
-
Modulation of tooth regeneration through opposing responses to Wnt and BMP signals in teleosts.Development. 2023 Dec 1;150(23):dev202168. doi: 10.1242/dev.202168. Epub 2023 Dec 7. Development. 2023. PMID: 38059590 Free PMC article.
-
Deficiency of Formyl Peptide Receptor 2 Retards Hair Regeneration by Modulating the Activation of Hair Follicle Stem Cells and Dermal Papilla Cells in Mice.Dev Reprod. 2021 Dec;25(4):279-291. doi: 10.12717/DR.2021.25.4.279. Epub 2021 Dec 31. Dev Reprod. 2021. PMID: 35141453 Free PMC article.
-
Computational modeling indicates that surface pressure can be reliably conveyed to tactile receptors even amidst changes in skin mechanics.J Neurophysiol. 2016 Jul 1;116(1):218-28. doi: 10.1152/jn.00624.2015. Epub 2016 Apr 20. J Neurophysiol. 2016. PMID: 27098029 Free PMC article.
References
-
- Andl T, Ahn K, Kairo A, Chu EY, Wine-Lee L, Reddy ST, Croft NJ, Cebra-Thomas JA, Metzger D, Chambon P, Lyons KM, Mishina Y, Seykora JT, Crenshaw EB, Millar SE. Epithelial Bmpr1a regulates differentiation and proliferation in postnatal hair follicles and is essential for tooth development. Development. 2004;131:2257–2268. - PubMed
-
- Blanpain C, Lowry WE, Geoghegan A, Polak L, Fuchs E. Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell. 2004;118:635–648. - PubMed
-
- Botchkarev VA, Botchkareva NV, Roth W, Nakamura M, Chen LH, Herzog W, Lindner G, Mcmahon JA, Peters C, Lauster R, Mcmahon AP, Paus R. Noggin is a mesenchymally derived stimulator of hair-follicle induction. Nat Cell Biol. 1999;1:158–164. - PubMed
-
- Botchkarev VA, Botchkareva NV, Nakamura M, Huber O, Funa K, Lauster R, Paus R, Gilchrest BA. Noggin is required for induction of the hair follicle growth phase in postnatal skin. FASEB J. 2001;5:2205–2214. - PubMed
-
- Botchkarev VA, Botchkareva NV, Huber O, Funa K, Gilchrest BA. Modulation of BMP signaling by noggin is required for induction of the secondary (non-tylotrich) hair follicles. J Invest Dermatol. 2002;118:3–10. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AR047364-07/AR/NIAMS NIH HHS/United States
- R01 AR042177-11/AR/NIAMS NIH HHS/United States
- R01 AR042177-13/AR/NIAMS NIH HHS/United States
- R01 AR042177-14/AR/NIAMS NIH HHS/United States
- R01 AR047364-06/AR/NIAMS NIH HHS/United States
- R01 AR042177-12S1/AR/NIAMS NIH HHS/United States
- R01 AR042177/AR/NIAMS NIH HHS/United States
- R01 AR047364/AR/NIAMS NIH HHS/United States
- R01 AR047364-04/AR/NIAMS NIH HHS/United States
- R01 AR042177-12/AR/NIAMS NIH HHS/United States
- R01 DE016320/DE/NIDCR NIH HHS/United States
- R01 DE016320-04/DE/NIDCR NIH HHS/United States
- R01 AR047364-05/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
