

Cofilin dissociates Arp2/3 complex and branches from actin filaments
- PMID: 19362000
- PMCID: PMC3711486
- DOI: 10.1016/j.cub.2009.02.060
Cofilin dissociates Arp2/3 complex and branches from actin filaments
Abstract
Background: Actin-based cellular motility requires spatially and temporally coordinated remodeling of a network of branched actin filaments. This study investigates how cofilin and Arp2/3 complex, two main players in the dendritic nucleation model, interact to produce sharp spatial transitions between densely branched filaments and long, unbranched filaments.
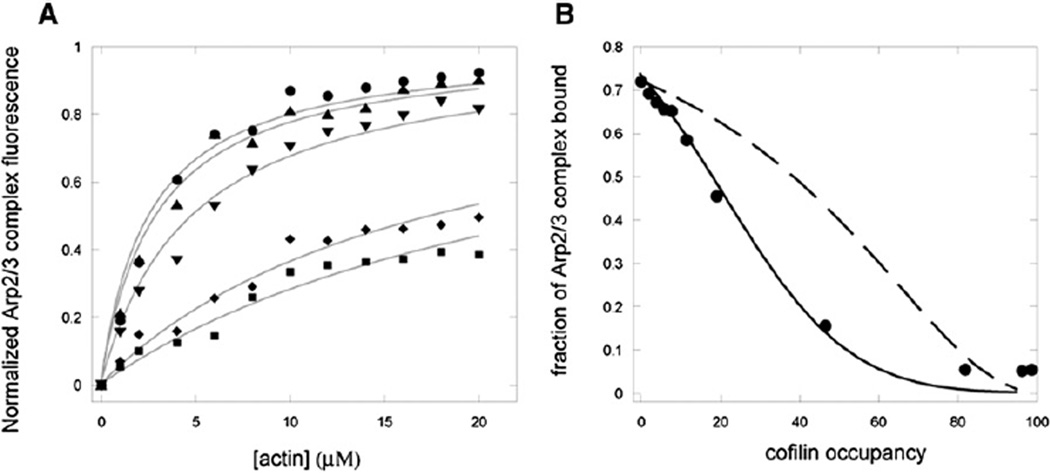
Results: We found that cofilin binding reduces both the affinity of actin filaments for Arp2/3 complex and the stability of branches. We used fluorescence spectroscopy to measure the kinetics of cofilin association with filaments and the resulting dissociation of Arp2/3 complex and TIRF microscopy to visualize filament severing and the loss of actin filament branches. Cofilin severs filaments optimally when few actin subunits are occupied but dissociates branches rapidly only at higher occupancies. Effective debranching is nevertheless achieved, as a result of cooperative binding and reduced affinity of Arp2/3 complex for the filament, at cofilin concentrations below those required for direct competition.
Conclusions: Cofilin rapidly dissociates Arp2/3 complex and branches by direct competition for binding sites on the actin filament and by propagation of structural changes in the actin filament that reduce affinity for Arp2/3 complex.
Figures



Similar articles
-
Interactions of ADF/cofilin, Arp2/3 complex, capping protein and profilin in remodeling of branched actin filament networks.Curr Biol. 2000 Oct 19;10(20):1273-82. doi: 10.1016/s0960-9822(00)00749-1. Curr Biol. 2000. PMID: 11069108
-
Mathematical modeling of endocytic actin patch kinetics in fission yeast: disassembly requires release of actin filament fragments.Mol Biol Cell. 2010 Aug 15;21(16):2905-15. doi: 10.1091/mbc.E10-06-0494. Epub 2010 Jun 29. Mol Biol Cell. 2010. PMID: 20587776 Free PMC article.
-
Arp2/3 complex and cofilin modulate binding of tropomyosin to branched actin networks.Curr Biol. 2015 Jun 15;25(12):1573-82. doi: 10.1016/j.cub.2015.04.038. Epub 2015 May 28. Curr Biol. 2015. PMID: 26028436 Free PMC article.
-
Biophysics of actin filament severing by cofilin.FEBS Lett. 2013 Apr 17;587(8):1215-9. doi: 10.1016/j.febslet.2013.01.062. Epub 2013 Feb 5. FEBS Lett. 2013. PMID: 23395798 Free PMC article. Review.
-
Cellular motility driven by assembly and disassembly of actin filaments.Cell. 2003 Feb 21;112(4):453-65. doi: 10.1016/s0092-8674(03)00120-x. Cell. 2003. PMID: 12600310 Review.
Cited by
-
Mechanisms of actin disassembly and turnover.J Cell Biol. 2023 Dec 4;222(12):e202309021. doi: 10.1083/jcb.202309021. Epub 2023 Nov 10. J Cell Biol. 2023. PMID: 37948068 Free PMC article. Review.
-
Direct observation of cortactin protecting Arp2/3-actin filament branch junctions from GMF-mediated destabilization.Eur J Cell Biol. 2024 Mar;103(1):151378. doi: 10.1016/j.ejcb.2023.151378. Epub 2023 Dec 5. Eur J Cell Biol. 2024. PMID: 38071835 Free PMC article.
-
GMF is a cofilin homolog that binds Arp2/3 complex to stimulate filament debranching and inhibit actin nucleation.Curr Biol. 2010 May 11;20(9):861-7. doi: 10.1016/j.cub.2010.03.026. Epub 2010 Apr 1. Curr Biol. 2010. PMID: 20362448 Free PMC article.
-
Unconcerted conformational changes in Arp2/3 complex integrate multiple activating signals to assemble functional actin networks.Curr Biol. 2022 Mar 14;32(5):975-987.e6. doi: 10.1016/j.cub.2022.01.004. Epub 2022 Jan 31. Curr Biol. 2022. PMID: 35090589 Free PMC article.
-
Rho GTPase signalling networks in cancer cell transendothelial migration.Vasc Biol. 2021 Sep 14;3(1):R77-R95. doi: 10.1530/VB-21-0008. eCollection 2021. Vasc Biol. 2021. PMID: 34738075 Free PMC article. Review.
References
-
- Chen H, Bernstein BW, Sneider JM, Boyle JA, Minamide LS, Bamburg JR. In vitro activity differences between proteins of the ADF/cofilin family define two distinct subgroups. Biochemistry. 2004;43:7127–7142. - PubMed
-
- Yeoh S, Pope B, Mannherz HG, Weeds A. Determining the differences in actin binding by human ADF and cofilin. J. Mol. Biol. 2002;315:911–925. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
