

Restriction of human polyomavirus BK virus DNA replication in murine cells and extracts
- PMID: 19297467
- PMCID: PMC2681987
- DOI: 10.1128/JVI.00300-09
Restriction of human polyomavirus BK virus DNA replication in murine cells and extracts
Abstract
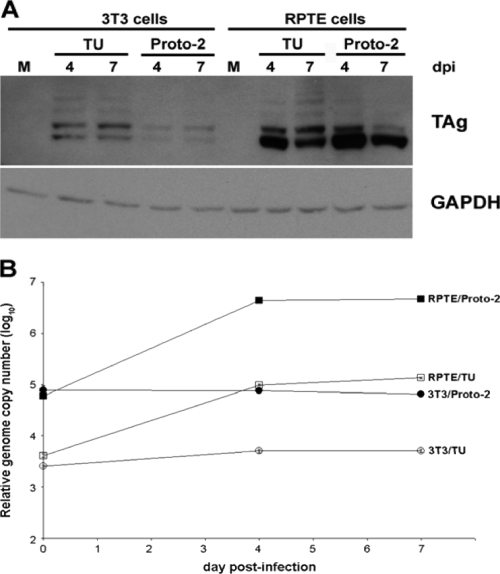
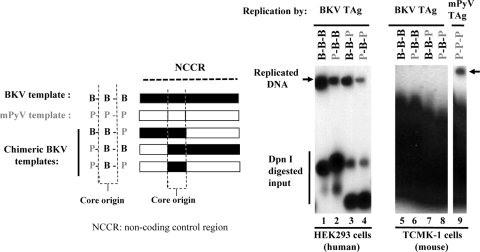
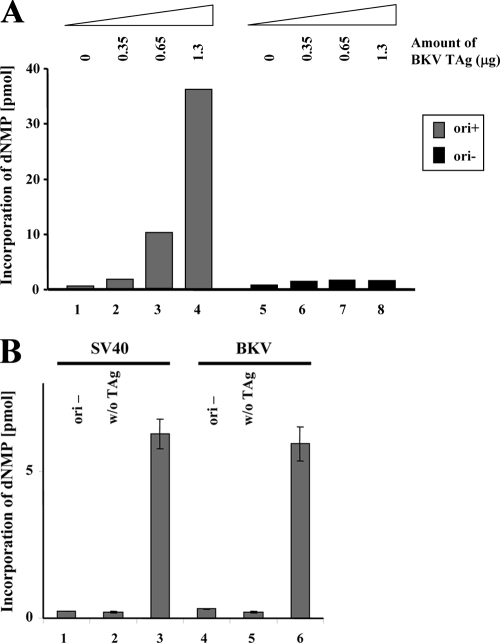
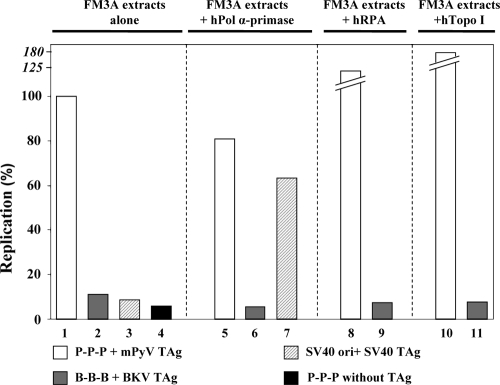
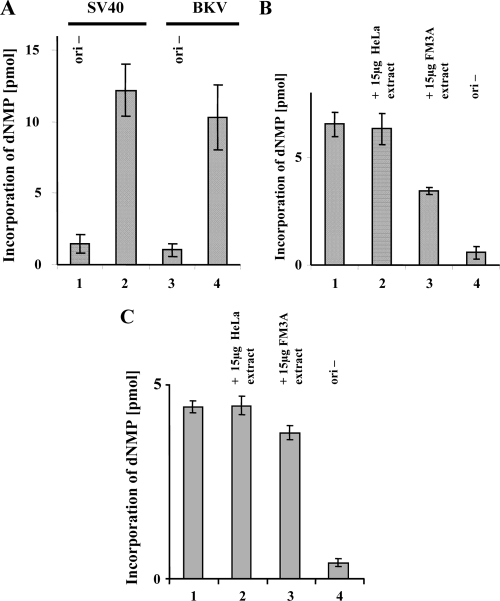
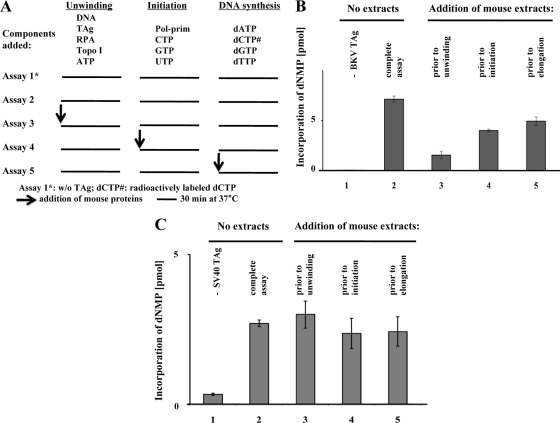
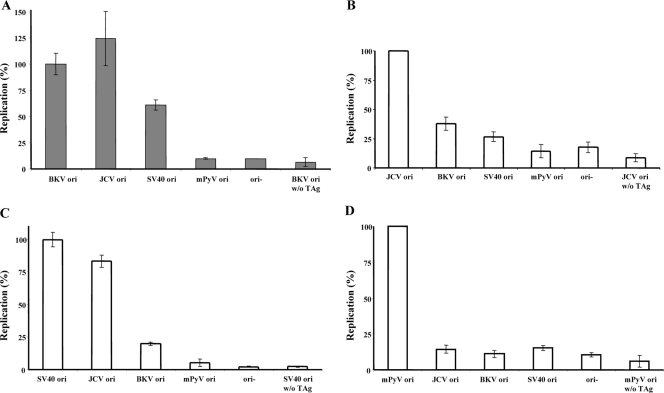
BK virus (BKV) causes persistent and asymptomatic infections in most humans and is the etiologic agent of polyomavirus-associated nephropathy (PVAN) and other pathologies. Unfortunately, there are no animal models with which to study activation of BKV replication in the human kidney and the accompanying PVAN. Here we report studies of the restriction of BKV replication in murine cells and extracts and the cause(s) of this restriction. Upon infection of murine cells, BKV expressed large T antigen (TAg), but viral DNA replication and progeny were not detected. Transfection of murine cells with BKV TAg expression vectors also caused TAg expression without accompanying DNA replication. Analysis of the replication of DNAs containing chimeric BKV and murine polyomavirus origins revealed the importance of BKV core origin sequences and TAg for DNA replication. A sensitive assay was developed with purified BKV TAg that supported TAg-dependent BKV DNA replication with human but not with murine cell extracts. Addition of human replication proteins, DNA polymerase alpha-primase, replication protein A, or topoisomerase I to the murine extracts with BKV TAg did not rescue viral DNA replication. Notably, addition of murine extracts to human extracts inhibited BKV TAg-dependent DNA replication at a step prior to or during unwinding of the viral origin. These findings and differences in replication specificity between BKV TAg and the TAgs of simian virus 40 (SV40) and JC virus (JCV) and their respective origins implicate features of the BKV TAg and origin distinct from SV40 and JCV in restriction of BKV replication in murine cells.
Figures
Similar articles
-
Stimulation of BK virus DNA replication by NFI family transcription factors.J Virol. 2012 Mar;86(6):3264-75. doi: 10.1128/JVI.06369-11. Epub 2011 Dec 28. J Virol. 2012. PMID: 22205750 Free PMC article.
-
Nucleotide sequence of the region encompassing the JC virus origin of DNA replication.J Virol. 1983 Apr;46(1):170-6. doi: 10.1128/JVI.46.1.170-176.1983. J Virol. 1983. PMID: 6298454 Free PMC article.
-
A cell-free replication system for human polyomavirus JC DNA.J Virol. 1997 Oct;71(10):7421-8. doi: 10.1128/JVI.71.10.7421-7428.1997. J Virol. 1997. PMID: 9311820 Free PMC article.
-
BK viral enhancer element and a human cellular homolog.Science. 1983 Nov 18;222(4625):749-55. doi: 10.1126/science.6314501. Science. 1983. PMID: 6314501 Review.
-
Regulation of gene expression in primate polyomaviruses.J Virol. 2009 Nov;83(21):10846-56. doi: 10.1128/JVI.00542-09. Epub 2009 Jul 29. J Virol. 2009. PMID: 19640999 Free PMC article. Review.
Cited by
-
Trinucleotide repeat expansions catalyzed by human cell-free extracts.Cell Res. 2013 Apr;23(4):565-72. doi: 10.1038/cr.2013.12. Epub 2013 Jan 22. Cell Res. 2013. PMID: 23337586 Free PMC article.
-
Inhibition of human BK polyomavirus replication by small noncoding RNAs.J Virol. 2011 Jul;85(14):6930-40. doi: 10.1128/JVI.00547-11. Epub 2011 May 4. J Virol. 2011. PMID: 21543481 Free PMC article.
-
Efficient propagation of archetype BK and JC polyomaviruses.Virology. 2012 Jan 20;422(2):235-41. doi: 10.1016/j.virol.2011.10.026. Epub 2011 Nov 17. Virology. 2012. PMID: 22099377 Free PMC article.
-
Alkoxylalkyl Esters of Nucleotide Analogs Inhibit Polyomavirus DNA Replication and Large T Antigen Activities.Antimicrob Agents Chemother. 2021 Feb 17;65(3):e01641-20. doi: 10.1128/AAC.01641-20. Print 2021 Feb 17. Antimicrob Agents Chemother. 2021. PMID: 33288638 Free PMC article.
-
Stimulation of BK virus DNA replication by NFI family transcription factors.J Virol. 2012 Mar;86(6):3264-75. doi: 10.1128/JVI.06369-11. Epub 2011 Dec 28. J Virol. 2012. PMID: 22205750 Free PMC article.
References
-
- Atencio, I. A., and L. P. Villarreal. 1994. Polyomavirus replicates in differentiating but not in proliferating tubules of adult mouse polycystic kidneys. Virology 20126-35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
