

Single-stranded DNA orchestrates an ATM-to-ATR switch at DNA breaks
- PMID: 19285939
- PMCID: PMC2675165
- DOI: 10.1016/j.molcel.2009.01.024
Single-stranded DNA orchestrates an ATM-to-ATR switch at DNA breaks
Abstract
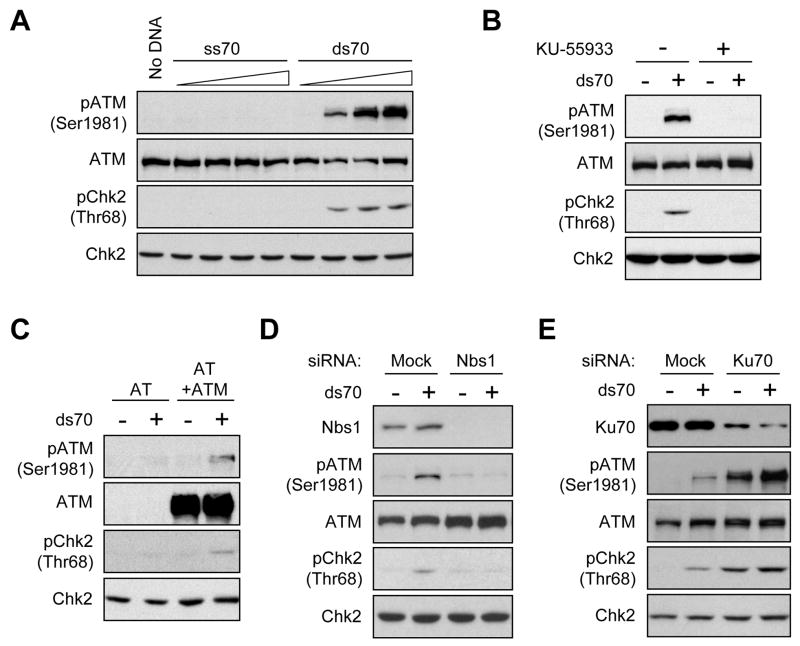
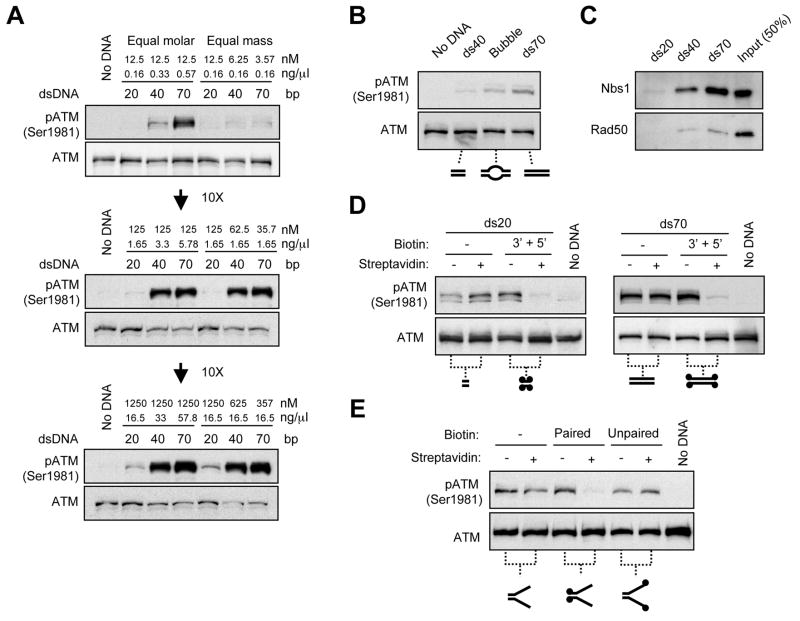
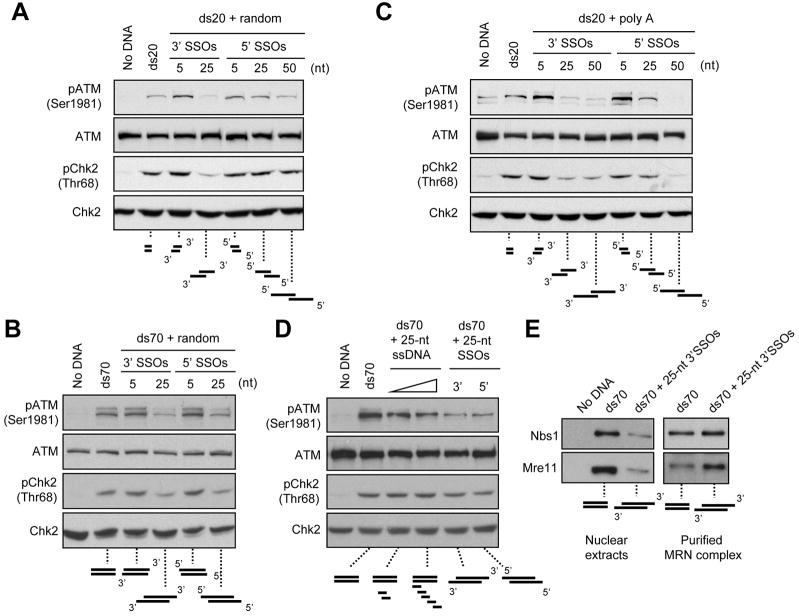
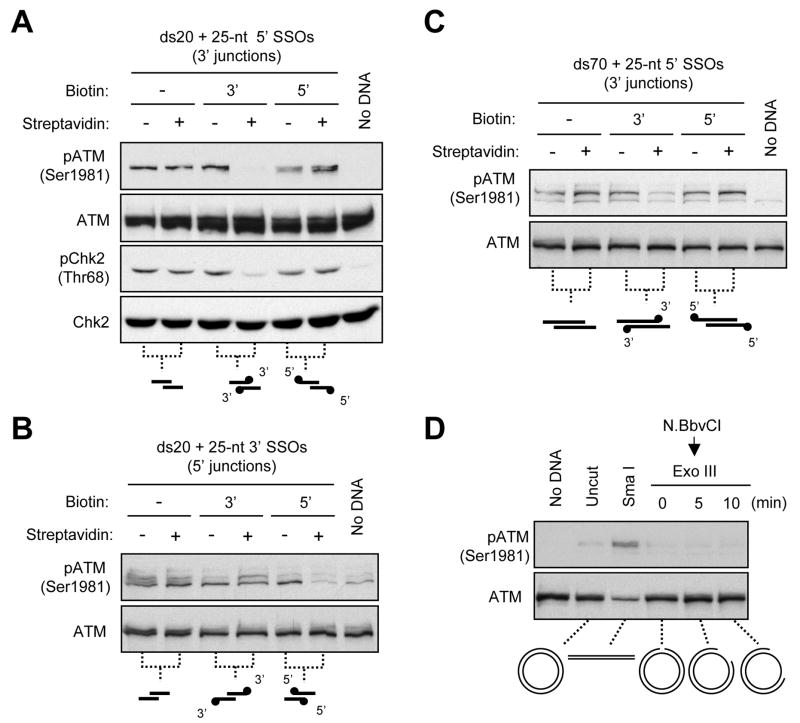
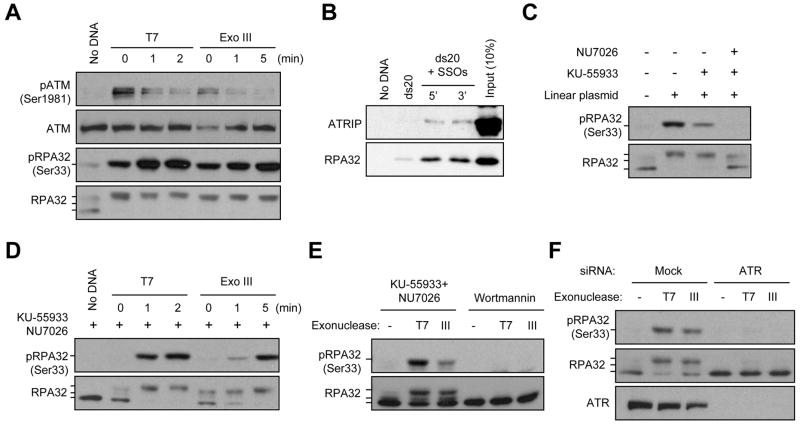
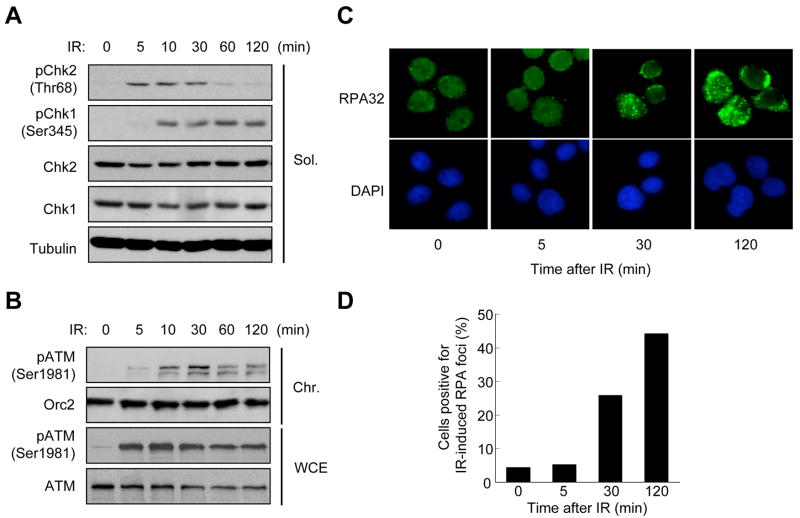
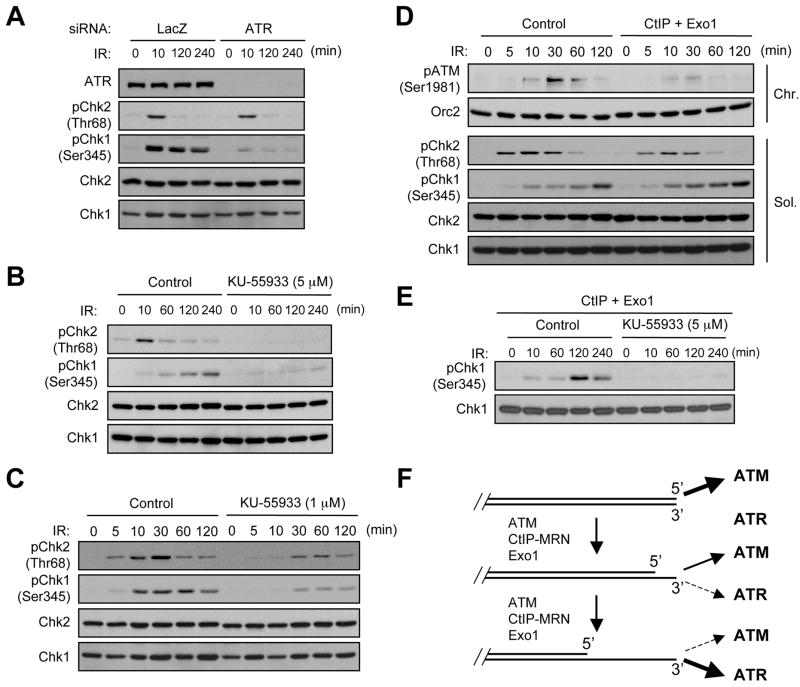
ATM and ATR are two master checkpoint kinases activated by double-stranded DNA breaks (DSBs). ATM is critical for the initial response and the subsequent ATR activation. Here we show that ATR activation is coupled with loss of ATM activation, an unexpected ATM-to-ATR switch during the biphasic DSB response. ATM is activated by DSBs with blunt ends or short single-stranded overhangs (SSOs). Surprisingly, the activation of ATM in the presence of SSOs, like that of ATR, relies on single- and double-stranded DNA junctions. In a length-dependent manner, SSOs attenuate ATM activation and potentiate ATR activation through a swap of DNA-damage sensors. Progressive resection of DSBs directly promotes the ATM-to-ATR switch in vitro. In cells, the ATM-to-ATR switch is driven by both ATM and the nucleases participating in DSB resection. Thus, single-stranded DNA orchestrates ATM and ATR to function in an orderly and reciprocal manner in two distinct phases of DSB response.
Figures
Comment in
-
Changing of the guard: how ATM hands off DNA double-strand break signaling to ATR.Mol Cell. 2009 Mar 27;33(6):672-4. doi: 10.1016/j.molcel.2009.03.004. Mol Cell. 2009. PMID: 19328060 Free PMC article.
Similar articles
-
Two distinct modes of ATR activation orchestrated by Rad17 and Nbs1.Cell Rep. 2013 May 30;3(5):1651-62. doi: 10.1016/j.celrep.2013.04.018. Epub 2013 May 16. Cell Rep. 2013. PMID: 23684611 Free PMC article.
-
ATM-dependent phosphorylation of the checkpoint clamp regulates repair pathways and maintains genomic stability.Cell Cycle. 2012 May 1;11(9):1796-803. doi: 10.4161/cc.20161. Epub 2012 May 1. Cell Cycle. 2012. PMID: 22453082 Free PMC article.
-
A role for the MRN complex in ATR activation via TOPBP1 recruitment.Mol Cell. 2013 Apr 11;50(1):116-22. doi: 10.1016/j.molcel.2013.03.006. Mol Cell. 2013. PMID: 23582259 Free PMC article.
-
Activation and regulation of ATM kinase activity in response to DNA double-strand breaks.Oncogene. 2007 Dec 10;26(56):7741-8. doi: 10.1038/sj.onc.1210872. Oncogene. 2007. PMID: 18066086 Review.
-
Mre11-Rad50-Nbs1 is a keystone complex connecting DNA repair machinery, double-strand break signaling, and the chromatin template.Biochem Cell Biol. 2007 Aug;85(4):509-20. doi: 10.1139/O07-069. Biochem Cell Biol. 2007. PMID: 17713585 Review.
Cited by
-
Induction of DNA damage by deguelin is mediated through reducing DNA repair genes in human non-small cell lung cancer NCI-H460 cells.Oncol Rep. 2012 Apr;27(4):959-64. doi: 10.3892/or.2012.1622. Epub 2012 Jan 4. Oncol Rep. 2012. PMID: 22227970 Free PMC article.
-
14-3-3 proteins restrain the Exo1 nuclease to prevent overresection.J Biol Chem. 2015 May 8;290(19):12300-12. doi: 10.1074/jbc.M115.644005. Epub 2015 Apr 1. J Biol Chem. 2015. PMID: 25833945 Free PMC article.
-
The DNA damage checkpoint: A tale from budding yeast.Front Genet. 2022 Sep 15;13:995163. doi: 10.3389/fgene.2022.995163. eCollection 2022. Front Genet. 2022. PMID: 36186482 Free PMC article. Review.
-
Telomere uncapping at the crossroad between cell cycle arrest and carcinogenesis.Mol Cell Oncol. 2014 Jul 28;1(1):e29901. doi: 10.4161/mco.29901. eCollection 2014. Mol Cell Oncol. 2014. PMID: 27308311 Free PMC article. Review.
-
Reconstitution of human claspin-mediated phosphorylation of Chk1 by the ATR (ataxia telangiectasia-mutated and rad3-related) checkpoint kinase.J Biol Chem. 2009 Nov 27;284(48):33107-14. doi: 10.1074/jbc.M109.064485. Epub 2009 Oct 14. J Biol Chem. 2009. PMID: 19828454 Free PMC article.
References
-
- Bakkenist CJ, Kastan MB. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 2003;421:499–506. - PubMed
-
- Bartkova J, Horejsi Z, Koed K, Kramer A, Tort F, Zieger K, Guldberg P, Sehested M, Nesland JM, Lukas C, et al. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature. 2005;434:864–870. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
