

Multifactorial regulation of a hox target gene
- PMID: 19282966
- PMCID: PMC2646128
- DOI: 10.1371/journal.pgen.1000412
Multifactorial regulation of a hox target gene
Erratum in
- PLoS Genet. 2009 Mar;5(3). doi: 10.1371/annotation/f89de76b-0e06-4086-994c-20d64c238a46. Stein, Sokrates M A [corrected to Stein, M A Sokrates]
Abstract
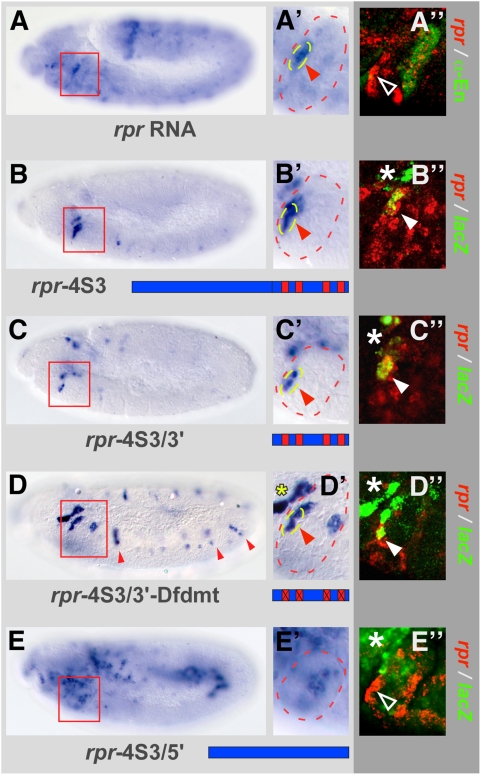
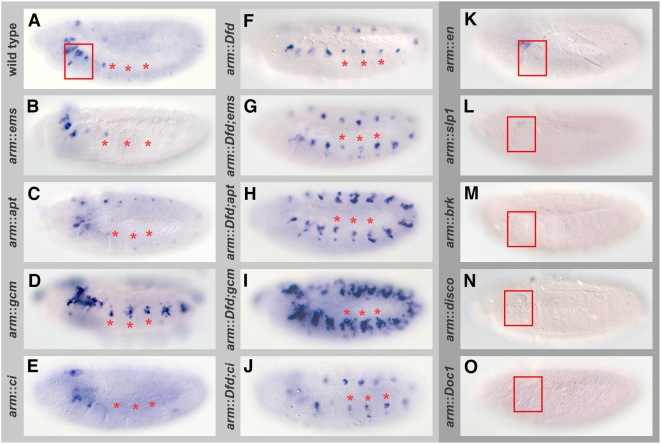
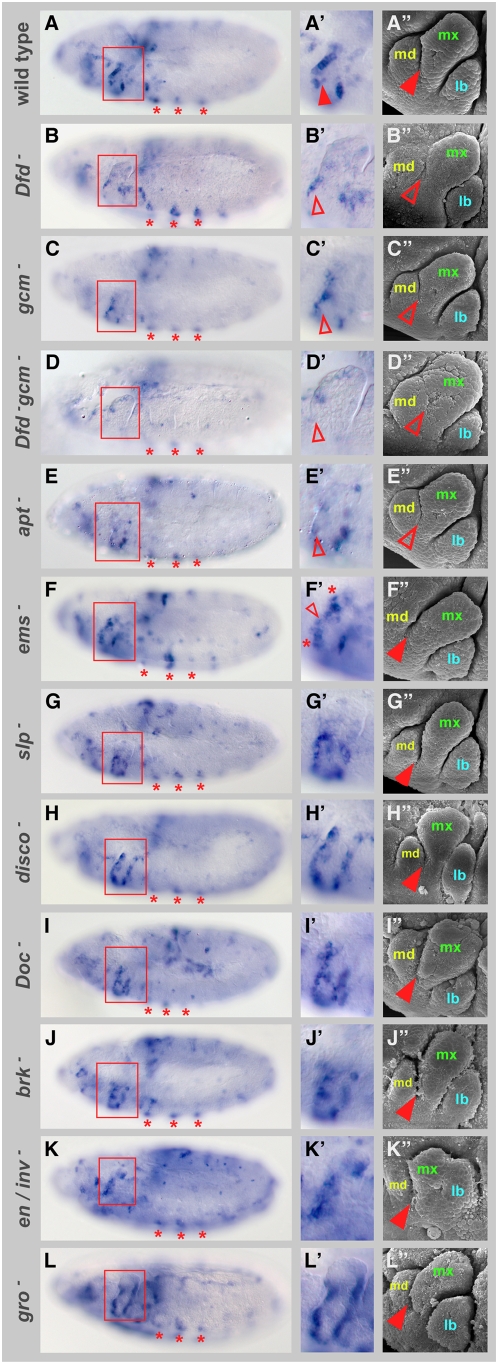
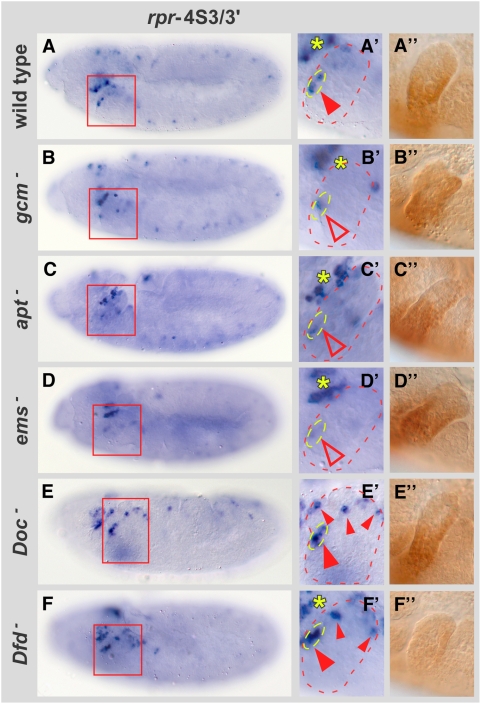
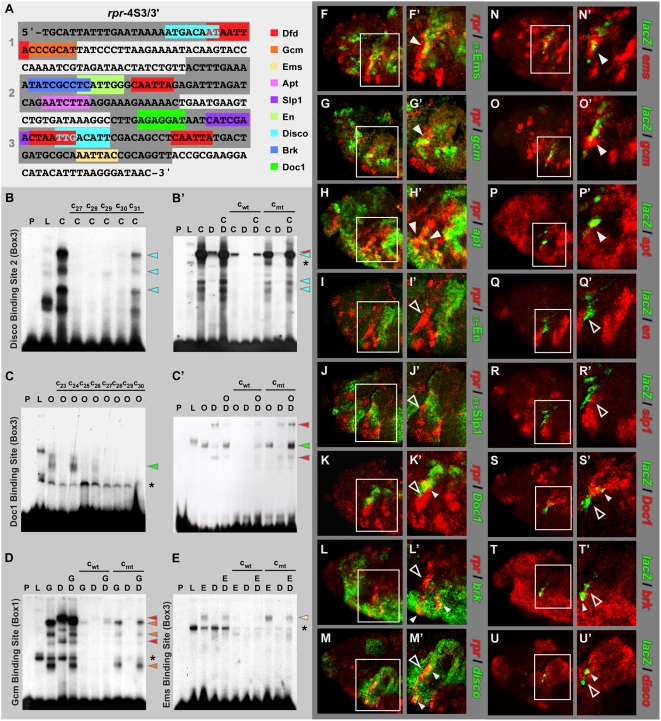
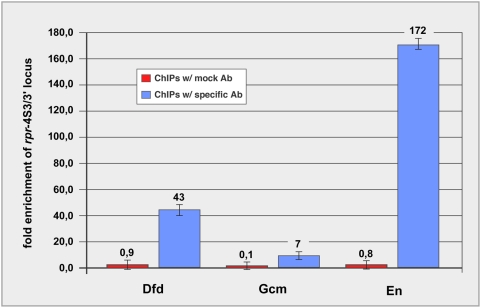
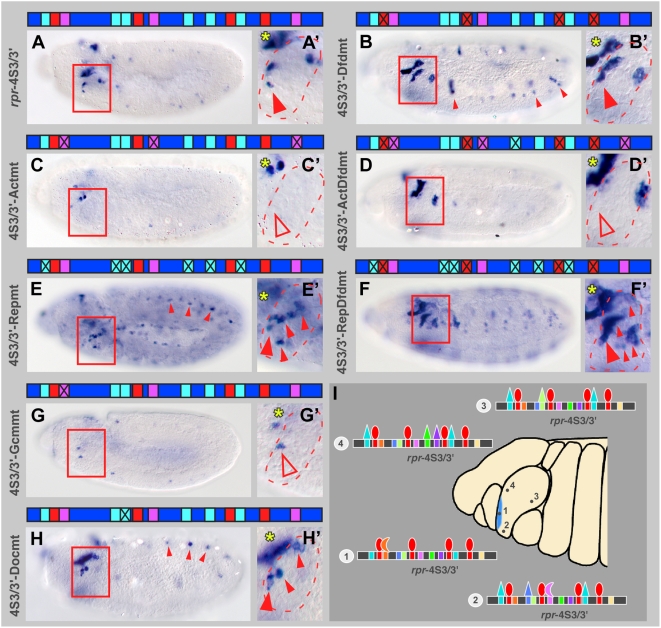
Hox proteins play fundamental roles in controlling morphogenetic diversity along the anterior-posterior body axis of animals by regulating distinct sets of target genes. Within their rather broad expression domains, individual Hox proteins control cell diversification and pattern formation and consequently target gene expression in a highly localized manner, sometimes even only in a single cell. To achieve this high-regulatory specificity, it has been postulated that Hox proteins co-operate with other transcription factors to activate or repress their target genes in a highly context-specific manner in vivo. However, only a few of these factors have been identified. Here, we analyze the regulation of the cell death gene reaper (rpr) by the Hox protein Deformed (Dfd) and suggest that local activation of rpr expression in the anterior part of the maxillary segment is achieved through a combinatorial interaction of Dfd with at least eight functionally diverse transcriptional regulators on a minimal enhancer. It follows that context-dependent combinations of Hox proteins and other transcription factors on small, modular Hox response elements (HREs) could be responsible for the proper spatio-temporal expression of Hox targets. Thus, a large number of transcription factors are likely to be directly involved in Hox target gene regulation in vivo.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The cis-regulatory code of Hox function in Drosophila.EMBO J. 2012 Aug 1;31(15):3323-33. doi: 10.1038/emboj.2012.179. Epub 2012 Jul 10. EMBO J. 2012. PMID: 22781127 Free PMC article.
-
Cellular analysis of newly identified Hox downstream genes in Drosophila.Eur J Cell Biol. 2010 Feb-Mar;89(2-3):273-8. doi: 10.1016/j.ejcb.2009.11.012. Epub 2009 Dec 16. Eur J Cell Biol. 2010. PMID: 20018403 Review.
-
The Drosophila Hox gene deformed sculpts head morphology via direct regulation of the apoptosis activator reaper.Cell. 2002 Aug 23;110(4):457-66. doi: 10.1016/s0092-8674(02)00871-1. Cell. 2002. PMID: 12202035
-
Specificity of the Hox member Deformed is determined by transcription factor levels and binding site affinities.Nat Commun. 2022 Aug 26;13(1):5037. doi: 10.1038/s41467-022-32408-8. Nat Commun. 2022. PMID: 36028502 Free PMC article.
-
Shaping segments: Hox gene function in the genomic age.Bioessays. 2008 Oct;30(10):965-79. doi: 10.1002/bies.20823. Bioessays. 2008. PMID: 18798525 Review.
Cited by
-
The cis-regulatory code of Hox function in Drosophila.EMBO J. 2012 Aug 1;31(15):3323-33. doi: 10.1038/emboj.2012.179. Epub 2012 Jul 10. EMBO J. 2012. PMID: 22781127 Free PMC article.
-
Activity of transcription factor JACKDAW is essential for SHR/SCR-dependent activation of SCARECROW and MAGPIE and is modulated by reciprocal interactions with MAGPIE, SCARECROW and SHORT ROOT.Plant Mol Biol. 2011 Nov;77(4-5):489-99. doi: 10.1007/s11103-011-9826-5. Epub 2011 Sep 21. Plant Mol Biol. 2011. PMID: 21935722
-
Disentangling the many layers of eukaryotic transcriptional regulation.Annu Rev Genet. 2012;46:43-68. doi: 10.1146/annurev-genet-110711-155437. Epub 2012 Aug 28. Annu Rev Genet. 2012. PMID: 22934649 Free PMC article. Review.
-
In silico characterization of the neural alpha tubulin gene promoter of the sea urchin embryo Paracentrotus lividus by phylogenetic footprinting.Mol Biol Rep. 2012 Mar;39(3):2633-44. doi: 10.1007/s11033-011-1016-7. Epub 2011 Jun 16. Mol Biol Rep. 2012. PMID: 21678058
-
Cell-type specific cis-regulatory networks: insights from Hox transcription factors.Fly (Austin). 2013 Jan-Mar;7(1):13-7. doi: 10.4161/fly.22939. Epub 2012 Dec 6. Fly (Austin). 2013. PMID: 23221502 Free PMC article.
References
-
- McGinnis W, Krumlauf R. Homeobox genes and axial patterning. Cell. 1992;68:283–302. - PubMed
-
- Mann RS, Morata G. The developmental and molecular biology of genes that subdivide the body of Drosophila. Annu Rev Cell Dev Biol. 2000;16:243–271. - PubMed
-
- Pearson JC, Lemons D, McGinnis W. Modulating Hox gene functions during animal body patterning. Nature Genetics Reviews. 2005;6:893–904. - PubMed
-
- Botas J. Control of morphogenesis and differentiation by HOM/Hox genes. Curr Opin Cell Biol. 1993;5:1015–1022. - PubMed
-
- Graba Y, Aragnol D, Pradel J. Drosophila Hox complex downstream targets and the function of homeotic genes. Bioessays. 1997;19:379–388. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
