

AtVPS45 is a positive regulator of the SYP41/SYP61/VTI12 SNARE complex involved in trafficking of vacuolar cargo
- PMID: 19251905
- PMCID: PMC2663731
- DOI: 10.1104/pp.108.134361
AtVPS45 is a positive regulator of the SYP41/SYP61/VTI12 SNARE complex involved in trafficking of vacuolar cargo
Abstract
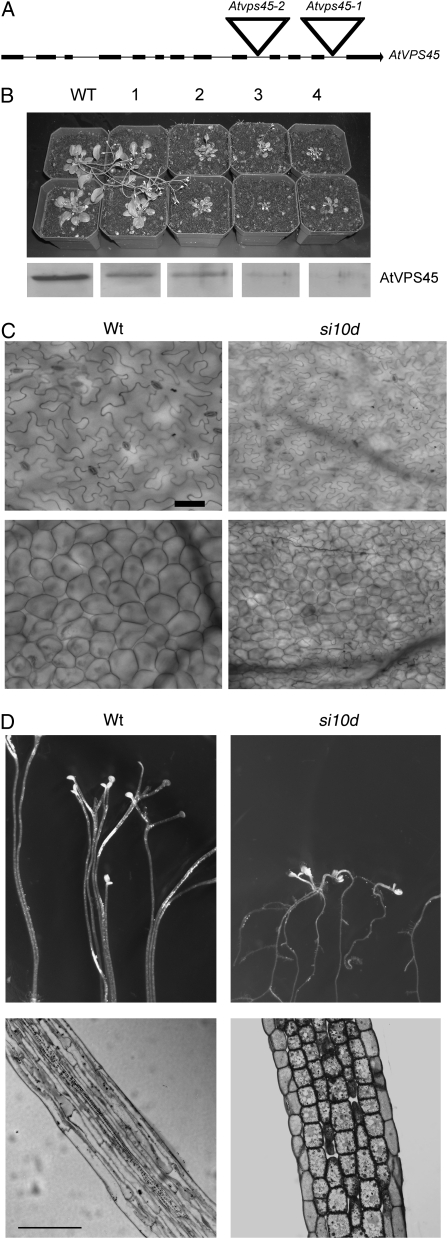
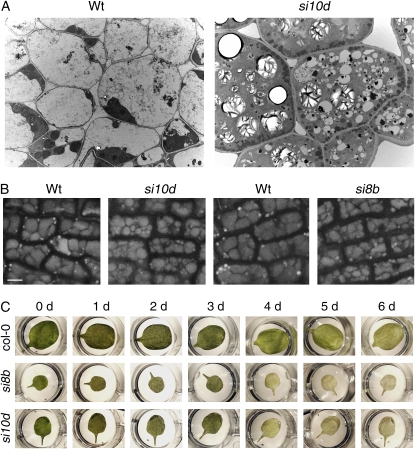
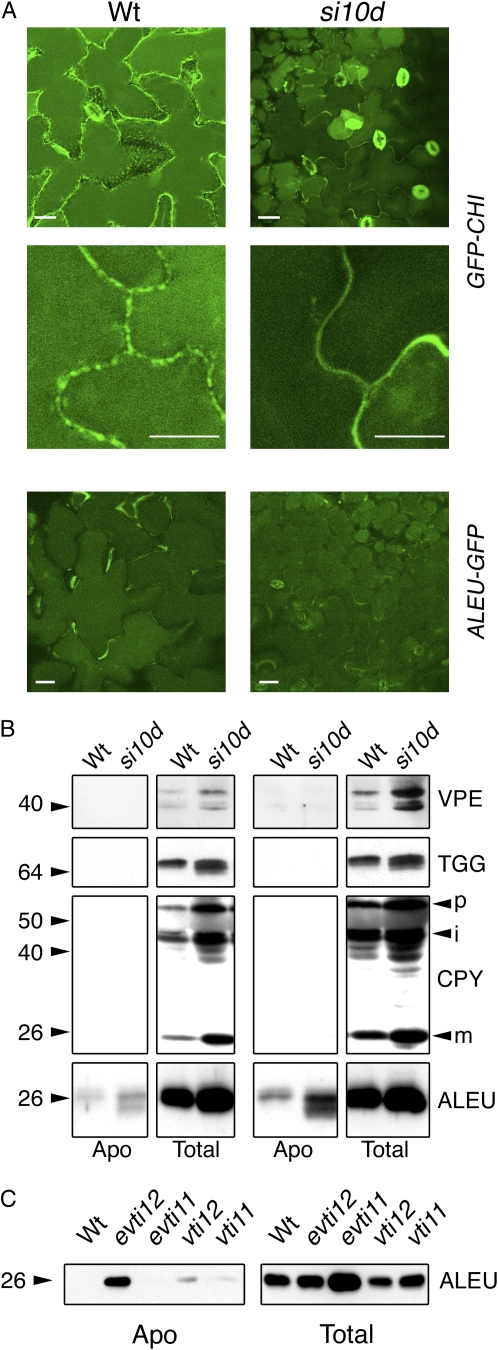
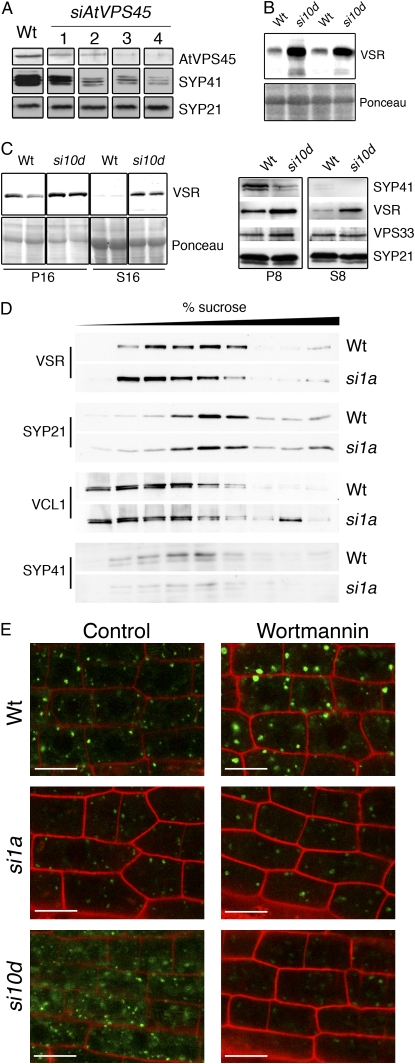
We report a functional characterization of AtVPS45 (for vacuolar protein sorting 45), a protein from the Sec1/Munc18 family in Arabidopsis (Arabidopsis thaliana) that interacts at the trans-Golgi network (TGN) with the SYP41/SYP61/VTI12 SNARE complex. A null allele of AtVPS45 was male gametophytic lethal, whereas stable RNA interference lines with reduced AtVPS45 protein levels had stunted growth but were viable and fertile. In the silenced lines, we observed defects in vacuole formation that correlated with a reduction in cell expansion and with autophagy-related defects in nutrient turnover. Moreover, transport of vacuolar cargo with carboxy-terminal vacuolar sorting determinants was blocked in the silenced lines, suggesting that AtVPS45 functions in vesicle trafficking to the vacuole. These trafficking defects are similar to those observed in vti12 mutants, supporting a functional relationship between AtVPS45 and VTI12. Consistent with this, we found a decrease in SYP41 protein levels coupled to the silencing of AtVPS45, pointing to instability and malfunction of the SYP41/SYP61/VTI12 SNARE complex in the absence of its cognate Sec1/Munc18 regulator. Based on its localization on the TGN, we hypothesized that AtVPS45 could be involved in membrane fusion of retrograde vesicles recycling vacuolar trafficking machinery. Indeed, in the AtVPS45-silenced plants, we found a striking alteration in the subcellular fractionation pattern of vacuolar sorting receptors, which are required for sorting of carboxy-terminal vacuolar sorting determinant-containing cargo. We propose that AtVPS45 is essential for recycling of the vacuolar sorting receptors back to the TGN and that blocking this step underlies the defects in vacuolar cargo trafficking observed in the silenced lines.
Figures

Similar articles
-
TNO1 is involved in salt tolerance and vacuolar trafficking in Arabidopsis.Plant Physiol. 2011 Jun;156(2):514-26. doi: 10.1104/pp.110.168963. Epub 2011 Apr 26. Plant Physiol. 2011. PMID: 21521696 Free PMC article.
-
Overexpression of trans-Golgi network t-SNAREs rescues vacuolar trafficking and TGN morphology defects in a putative tethering factor mutant.Plant J. 2019 Aug;99(4):703-716. doi: 10.1111/tpj.14353. Epub 2019 Jun 4. Plant J. 2019. PMID: 31009161
-
AtVPS45 complex formation at the trans-Golgi network.Mol Biol Cell. 2000 Jul;11(7):2251-65. doi: 10.1091/mbc.11.7.2251. Mol Biol Cell. 2000. PMID: 10888666 Free PMC article.
-
The riddle of the plant vacuolar sorting receptors.Protoplasma. 2005 Dec;226(3-4):103-8. doi: 10.1007/s00709-005-0117-3. Epub 2005 Dec 12. Protoplasma. 2005. PMID: 16333569 Review.
-
Lysosomal and vacuolar sorting: not so different after all!Biochem Soc Trans. 2016 Jun 15;44(3):891-7. doi: 10.1042/BST20160050. Biochem Soc Trans. 2016. PMID: 27284057 Free PMC article. Review.
Cited by
-
Arabidopsis Sec1/Munc18 protein SEC11 is a competitive and dynamic modulator of SNARE binding and SYP121-dependent vesicle traffic.Plant Cell. 2013 Apr;25(4):1368-82. doi: 10.1105/tpc.112.108506. Epub 2013 Apr 9. Plant Cell. 2013. PMID: 23572542 Free PMC article.
-
Proteomic analysis of insulin secretory granules in INS-1 cells by protein correlation profiling.Biophys Rep. 2018;4(6):329-338. doi: 10.1007/s41048-018-0061-3. Epub 2018 Aug 29. Biophys Rep. 2018. PMID: 30596141 Free PMC article.
-
Membrane traffic and fusion at post-Golgi compartments.Front Plant Sci. 2012 Jan 4;2:111. doi: 10.3389/fpls.2011.00111. eCollection 2011. Front Plant Sci. 2012. PMID: 22645561 Free PMC article.
-
The deubiquitinating enzyme AMSH3 is required for intracellular trafficking and vacuole biogenesis in Arabidopsis thaliana.Plant Cell. 2010 Jun;22(6):1826-37. doi: 10.1105/tpc.110.075952. Epub 2010 Jun 11. Plant Cell. 2010. PMID: 20543027 Free PMC article.
-
Cell polarity and patterning by PIN trafficking through early endosomal compartments in Arabidopsis thaliana.PLoS Genet. 2013 May;9(5):e1003540. doi: 10.1371/journal.pgen.1003540. Epub 2013 May 30. PLoS Genet. 2013. PMID: 23737757 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
