

Functional screen reveals SARS coronavirus nonstructural protein nsp14 as a novel cap N7 methyltransferase
- PMID: 19208801
- PMCID: PMC2651275
- DOI: 10.1073/pnas.0808790106
Functional screen reveals SARS coronavirus nonstructural protein nsp14 as a novel cap N7 methyltransferase
Abstract
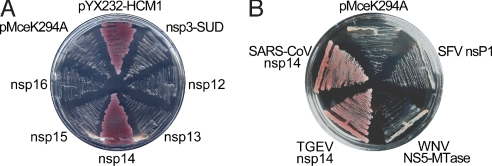
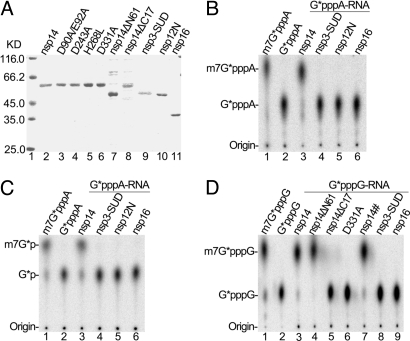
The N7-methylguanosine (m7G) cap is the defining structural feature of eukaryotic mRNAs. Most eukaryotic viruses that replicate in the cytoplasm, including coronaviruses, have evolved strategies to cap their RNAs. In this report, we used a yeast genetic system to functionally screen for the cap-forming enzymes encoded by severe acute respiratory syndrome (SARS) coronavirus and identified the nonstructural protein (nsp) 14 of SARS coronavirus as a (guanine-N7)-methyltransferase (N7-MTase) in vivo in yeast cells and in vitro using purified enzymes and RNA substrates. Interestingly, coronavirus nsp14 was previously characterized as a 3'-to-5' exoribonuclease, and by mutational analysis, we mapped the N7-MTase domain to the carboxy-terminal part of nsp14 that shows features conserved with cellular N7-MTase in structure-based sequence alignment. The exoribonuclease active site was dispensable but the exoribonuclease domain was required for N7-MTase activity. Such combination of the 2 functional domains in coronavirus nsp14 suggests that it may represent a novel form of RNA-processing enzymes. Mutational analysis in a replicon system showed that the N7-MTase activity was important for SARS virus replication/transcription and can thus be used as an attractive drug target to develop antivirals for control of coronaviruses including the deadly SARS virus. Furthermore, the observation that the N7-MTase of RNA life could function in lieu of that in DNA life provides interesting evolutionary insight and practical possibilities in antiviral drug screening.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Structure-function analysis of severe acute respiratory syndrome coronavirus RNA cap guanine-N7-methyltransferase.J Virol. 2013 Jun;87(11):6296-305. doi: 10.1128/JVI.00061-13. Epub 2013 Mar 27. J Virol. 2013. PMID: 23536667 Free PMC article.
-
Mutagenesis of S-Adenosyl-l-Methionine-Binding Residues in Coronavirus nsp14 N7-Methyltransferase Demonstrates Differing Requirements for Genome Translation and Resistance to Innate Immunity.J Virol. 2016 Jul 27;90(16):7248-7256. doi: 10.1128/JVI.00542-16. Print 2016 Aug 15. J Virol. 2016. PMID: 27252528 Free PMC article.
-
Characterization of the guanine-N7 methyltransferase activity of coronavirus nsp14 on nucleotide GTP.Virus Res. 2013 Sep;176(1-2):45-52. doi: 10.1016/j.virusres.2013.05.001. Epub 2013 May 20. Virus Res. 2013. PMID: 23702198 Free PMC article.
-
SARS-CoV ORF1b-encoded nonstructural proteins 12-16: replicative enzymes as antiviral targets.Antiviral Res. 2014 Jan;101:122-30. doi: 10.1016/j.antiviral.2013.11.006. Epub 2013 Nov 20. Antiviral Res. 2014. PMID: 24269475 Free PMC article. Review.
-
The Nonstructural Proteins Directing Coronavirus RNA Synthesis and Processing.Adv Virus Res. 2016;96:59-126. doi: 10.1016/bs.aivir.2016.08.008. Epub 2016 Sep 14. Adv Virus Res. 2016. PMID: 27712628 Free PMC article. Review.
Cited by
-
Development and characterization of reverse genetics systems of feline infectious peritonitis virus for antiviral research.Vet Res. 2024 Sep 27;55(1):124. doi: 10.1186/s13567-024-01373-z. Vet Res. 2024. PMID: 39334482 Free PMC article.
-
Comparative Atlas of SARS-CoV-2 Substitution Mutations: A Focus on Iranian Strains Amidst Global Trends.Viruses. 2024 Aug 20;16(8):1331. doi: 10.3390/v16081331. Viruses. 2024. PMID: 39205305 Free PMC article.
-
Long COVID Definition, Symptoms, Risk Factors, Epidemiology and Autoimmunity: A Narrative Review.Am J Med Open. 2024 Feb 14;11:100068. doi: 10.1016/j.ajmo.2024.100068. eCollection 2024 Jun. Am J Med Open. 2024. PMID: 39034937 Free PMC article.
-
Structural and functional insights into the 2'-O-methyltransferase of SARS-CoV-2.Virol Sin. 2024 Aug;39(4):619-631. doi: 10.1016/j.virs.2024.07.001. Epub 2024 Jul 3. Virol Sin. 2024. PMID: 38969340 Free PMC article.
-
Natural evidence of coronaviral 2'-O-methyltransferase activity affecting viral pathogenesis via improved substrate RNA binding.Signal Transduct Target Ther. 2024 May 29;9(1):140. doi: 10.1038/s41392-024-01860-x. Signal Transduct Target Ther. 2024. PMID: 38811528 Free PMC article.
References
-
- Ogino T, Banerjee AK. Unconventional mechanism of mRNA capping by the RNA-dependent RNA polymerase of vesicular stomatitis virus. Mol Cell. 2007;25:85–97. - PubMed
-
- Drosten C, et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N Engl J Med. 2003;348:1967–1976. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
