

Review
doi: 10.1016/j.ceb.2008.12.002.
Epub 2009 Jan 27.
Walking the walk: how kinesin and dynein coordinate their steps
Affiliations
- PMID: 19179063
- PMCID: PMC2668149
- DOI: 10.1016/j.ceb.2008.12.002
Item in Clipboard
Review
Walking the walk: how kinesin and dynein coordinate their steps
Curr Opin Cell Biol.
2009 Feb.
Abstract
Molecular motors drive key biological processes such as cell division, intracellular organelle transport, and sperm propulsion and defects in motor function can give rise to various human diseases. Two dimeric microtubule-based motor proteins, kinesin-1 and cytoplasmic dynein can take over one hundred steps without detaching from the track. In this review, we discuss how these processive motors coordinate the activities of their two identical motor domains so that they can walk along microtubules.
Figures
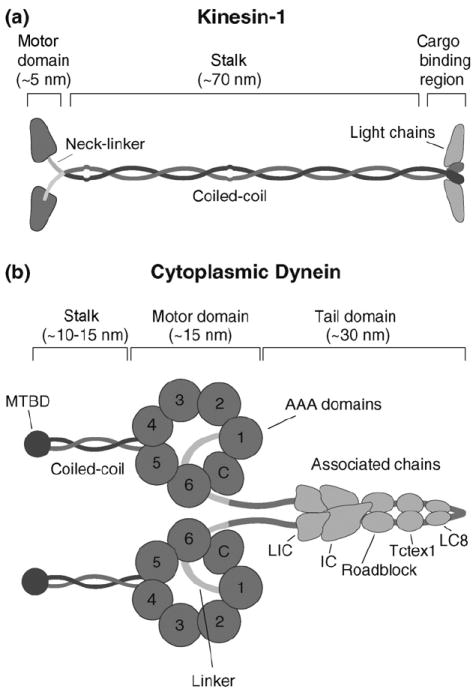
(a) Kinesin structure. Kinesin is composed of two identical heavy chains and two light chains. The heavy chain contains an N-terminal globular motor domain that possesses catalytic and MT-binding activity, a neck-linker element that connects the motor domain to the common coiled-coil dimerization domain, and a C-terminal light chain and cargo-binding region. (b) Cartoon representation of dynein. Dynein is composed of two identical heavy chains and several associated chains. The heavy chain forms a C-terminal catalytic motor domain that consists of multiple AAA ATP-binding sites (1–4) and a coiled-coil stalk with the microtubule-binding domain (MTBD) at its tip; AAA domains 5 and 6 do not contain sequences associated with nucleotide binding and the C-terminus (C) does not contain sequences characteristic for AAA proteins. The motor and dimerization domains are joined by a linker element. Multiple associated chains bind to dynein’s tail domain (LIC, light intermediate chain; IC, intermediate chain; Roadblock, Tctex1 and LC8, light chains).
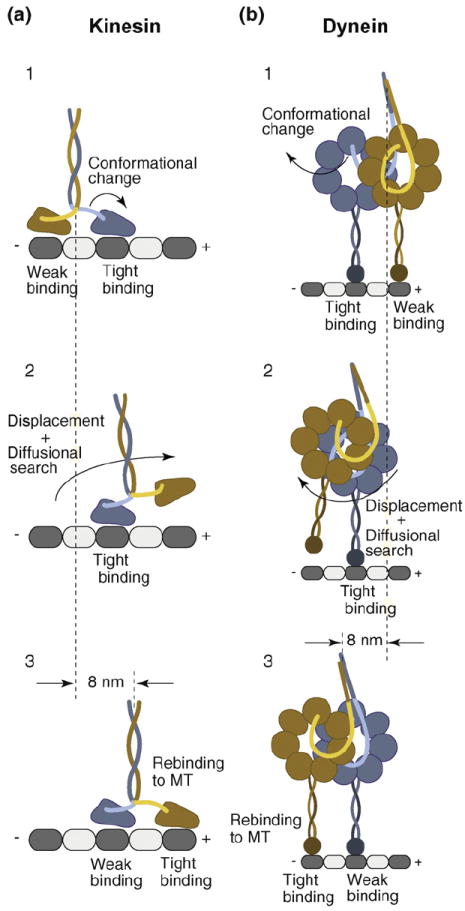
(a) Consensus stepping sequence of kinesin (for a detailed description, see text). A nucleotide-driven conformational change in the tightly MT-interacting front head displaces the weakly MT-interacting rear head toward the MT plus-end, biasing its diffusional search and rebinding to the next available MT-binding site in front of its partner head. While the rear head undergoes a 16 nm displacement, kinesin’s center-of-mass advances 8 nm. (b) Possible dynein stepping sequence. Dynein’s head domains are MT-bound with partially overlapping AAA rings, aligned parallel to the long MT axis. A nucleotide-dependent conformational change of the linker element in the tightly MT-binding front head displaces the weakly MT-interacting partner head toward the MT minus-end (opposite to the direction of kinesin movement). The displaced head then undergoes a rapid diffusional search and rebinds to the MT, resulting in a center-of-mass movement of 8 nm. Although dynein takes predominantly 8 nm steps, it has a considerable diffusional component to its step, resulting in different sized center-of-mass steps (4–24 nm).
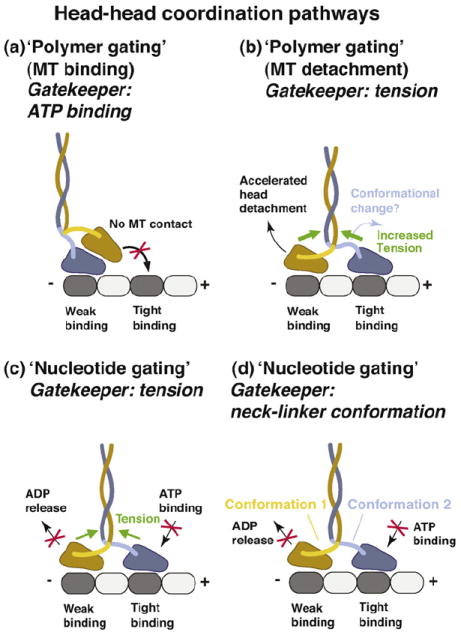
(a) In this tension-independent, ‘polymer gate’ pathway, the tethered head is detached from the MT and parked against the MT-bound head toward the MT plus-end. This interaction prevents the tethered head from binding to tubulin. The gate is opened when ATP binds to the MT-bound head (the ‘gatekeeper’ is ATP binding). (b) Internal tension might affect the MT-affinity of the head domains (‘polymer gating’). In this pathway, forward tension on the rear head is suggested to weaken its MT-affinity and promote its detachment. Acceleration of rear head detachment might be enhanced by the initiation of neck-linker docking in the front head (the ATP-driven conformation change). However, to be consistent with models in which the front head cannot bind ATP until the rear head detaches [50], the ADP-bound rear head must be sufficiently mobile to relieve the block on the front head. (c) Internal tension (the ‘gatekeeper’), derived from the simultaneous binding of both motor domains to the MT, controls the binding/release of nucleotides (‘nucleotide gating’). Backward tension on the front head transmitted via the neck-linker elements is suggested to prevent ATP binding, while the forward tension on the rear head is proposed to inhibit ADP release, thus helping to keep both heads out-of-phase. (d) Neck-linker conformation-based mechanism. In this pathway, the neck-linker elements might act as ‘control levers’ to regulate the binding/release of nucleotides allosterically. The backward-pointing/backward-docked orientation in the front head might prevent ATP binding and the forward-pointing/ forward-docked orientation in the trailing head might inhibit ADP release. Although tension might assist in creating and/or maintaining these conformations, the neck-linker conformation-based mechanism discussed here envisions the neck linkers as active regulatory elements, in contrast to the tension-based gating mechanisms described in parts (b) and (c) of this figure, in which the neck-linker elements serve as passive intermediaries that relay tension to the nucleotide and polymer binding sites.
Similar articles
-
Kinesin-1, -2, and -3 motors use family-specific mechanochemical strategies to effectively compete with dynein during bidirectional transport.Elife. 2022 Sep 20;11:e82228. doi: 10.7554/eLife.82228. Elife. 2022. PMID: 36125250 Free PMC article.
-
Intraflagellar transport dynein is autoinhibited by trapping of its mechanical and track-binding elements.Nat Struct Mol Biol. 2017 May;24(5):461-468. doi: 10.1038/nsmb.3391. Epub 2017 Apr 10. Nat Struct Mol Biol. 2017. PMID: 28394326 Free PMC article.
-
The types and numbers of kinesins and dyneins transporting endocytic cargoes modulate their motility and response to tau.J Biol Chem. 2024 Jun;300(6):107323. doi: 10.1016/j.jbc.2024.107323. Epub 2024 Apr 25. J Biol Chem. 2024. PMID: 38677516 Free PMC article.
-
How Dynein Moves Along Microtubules.Trends Biochem Sci. 2016 Jan;41(1):94-105. doi: 10.1016/j.tibs.2015.11.004. Epub 2015 Dec 9. Trends Biochem Sci. 2016. PMID: 26678005 Free PMC article. Review.
-
Motor Skills: Recruitment of Kinesins, Myosins and Dynein during Assembly and Egress of Alphaherpesviruses.Viruses. 2021 Aug 17;13(8):1622. doi: 10.3390/v13081622. Viruses. 2021. PMID: 34452486 Free PMC article. Review.
Cited by
-
Endolysosomal vesicles at the center of B cell activation.J Cell Biol. 2024 Mar 4;223(3):e202307047. doi: 10.1083/jcb.202307047. Epub 2024 Feb 2. J Cell Biol. 2024. PMID: 38305771 Free PMC article. Review.
-
Key residues on microtubule responsible for activation of kinesin ATPase.EMBO J. 2010 Apr 7;29(7):1167-75. doi: 10.1038/emboj.2010.25. Epub 2010 Mar 11. EMBO J. 2010. PMID: 20224548 Free PMC article.
-
Phosphorylation-independent dual-site binding of the FHA domain of KIF13 mediates phosphoinositide transport via centaurin alpha1.Proc Natl Acad Sci U S A. 2010 Nov 23;107(47):20346-51. doi: 10.1073/pnas.1009008107. Epub 2010 Nov 5. Proc Natl Acad Sci U S A. 2010. PMID: 21057110 Free PMC article.
-
The auto-inhibitory domain and ATP-independent microtubule-binding region of Kinesin heavy chain are major functional domains for transport in the Drosophila germline.Development. 2014 Jan;141(1):176-86. doi: 10.1242/dev.097592. Epub 2013 Nov 20. Development. 2014. PMID: 24257625 Free PMC article.
-
These motors were made for walking.Protein Sci. 2020 Aug;29(8):1707-1723. doi: 10.1002/pro.3895. Epub 2020 Jun 26. Protein Sci. 2020. PMID: 32472639 Free PMC article. Review.
References
-
- Vale RD. The molecular motor toolbox for intracellular transport. Cell. 2003;112:467–480. - PubMed
-
- Vallee RB, Williams JC, Varma D, Barnhart LE. Dynein: an ancient motor protein involved in multiple modes of transport. J Neurobiol. 2004;58:189–200. - PubMed
-
- Howard J, Hudspeth AJ, Vale RD. Movement of microtubules by single kinesin molecules. Nature. 1989;342:154–158. - PubMed
-
- Block SM, Goldstein LS, Schnapp BJ. Bead movement by single kinesin molecules studied with optical tweezers. Nature. 1990;348:348–352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
