

Primary microRNA transcripts are processed co-transcriptionally
- PMID: 19172742
- PMCID: PMC6952270
- DOI: 10.1038/nsmb.1475
Primary microRNA transcripts are processed co-transcriptionally
Abstract
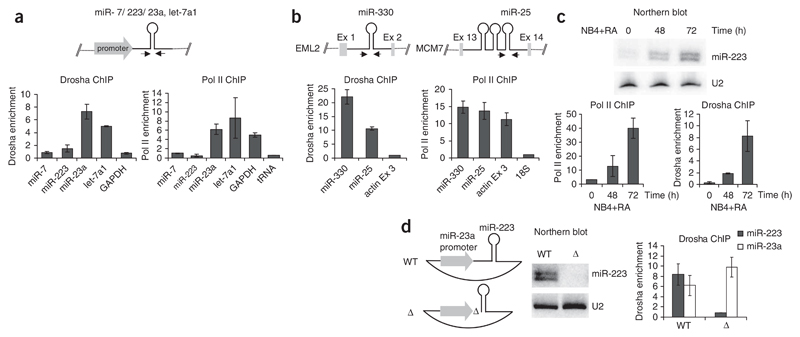
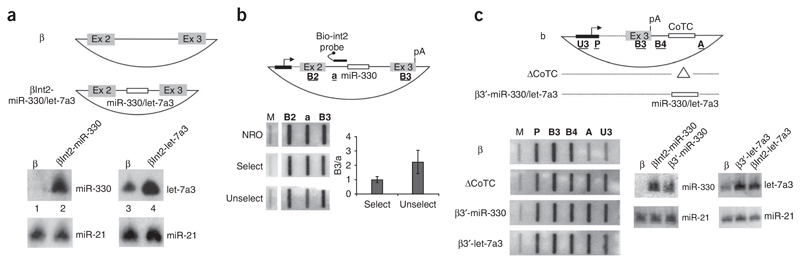
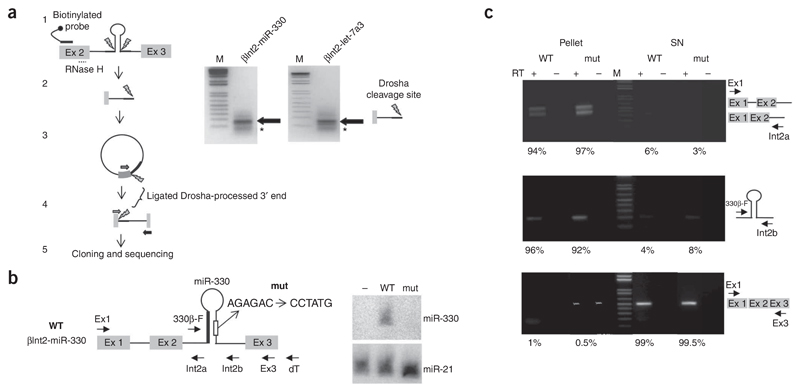
microRNAs (miRNAs) are generated from long primary (pri-) RNA polymerase II (Pol II)-derived transcripts by two RNase III processing reactions: Drosha cleavage of nuclear pri-miRNAs and Dicer cleavage of cytoplasmic pre-miRNAs. Here we show that Drosha cleavage occurs during transcription acting on both independently transcribed and intron-encoded miRNAs. We also show that both 5'-3' and 3'-5' exonucleases associate with the sites where co-transcriptional Drosha cleavage occurs, promoting intron degradation before splicing. We finally demonstrate that miRNAs can also derive from 3' flanking transcripts of Pol II genes. Our results demonstrate that multiple miRNA-containing transcripts are co-transcriptionally cleaved during their synthesis and suggest that exonucleolytic degradation from Drosha cleavage sites in pre-mRNAs may influence the splicing and maturation of numerous mRNAs.
Figures


Similar articles
-
HP1BP3, a Chromatin Retention Factor for Co-transcriptional MicroRNA Processing.Mol Cell. 2016 Aug 4;63(3):420-32. doi: 10.1016/j.molcel.2016.06.014. Epub 2016 Jul 14. Mol Cell. 2016. PMID: 27425409 Free PMC article.
-
Coupled RNA processing and transcription of intergenic primary microRNAs.Mol Cell Biol. 2009 Oct;29(20):5632-8. doi: 10.1128/MCB.00664-09. Epub 2009 Aug 10. Mol Cell Biol. 2009. PMID: 19667074 Free PMC article.
-
A central role for the primary microRNA stem in guiding the position and efficiency of Drosha processing of a viral pri-miRNA.RNA. 2014 Jul;20(7):1068-77. doi: 10.1261/rna.044537.114. Epub 2014 May 22. RNA. 2014. PMID: 24854622 Free PMC article.
-
The role of the precursor structure in the biogenesis of microRNA.Cell Mol Life Sci. 2011 Sep;68(17):2859-71. doi: 10.1007/s00018-011-0726-2. Epub 2011 May 24. Cell Mol Life Sci. 2011. PMID: 21607569 Free PMC article. Review.
-
Mirtrons: microRNA biogenesis via splicing.Biochimie. 2011 Nov;93(11):1897-904. doi: 10.1016/j.biochi.2011.06.017. Epub 2011 Jun 21. Biochimie. 2011. PMID: 21712066 Free PMC article. Review.
Cited by
-
Ensemble analysis of primary microRNA structure reveals an extensive capacity to deform near the Drosha cleavage site.Biochemistry. 2013 Feb 5;52(5):795-807. doi: 10.1021/bi301452a. Epub 2013 Jan 18. Biochemistry. 2013. PMID: 23305493 Free PMC article.
-
Transcripts for combined synthetic microRNA and gene delivery.Mol Biosyst. 2013 Jul;9(7):1919-25. doi: 10.1039/c3mb70043g. Epub 2013 Apr 12. Mol Biosyst. 2013. PMID: 23579254 Free PMC article.
-
Genome-wide transcript profiling reveals novel breast cancer-associated intronic sense RNAs.PLoS One. 2015 Mar 23;10(3):e0120296. doi: 10.1371/journal.pone.0120296. eCollection 2015. PLoS One. 2015. PMID: 25798919 Free PMC article.
-
MicroRNA biogenesis: regulating the regulators.Crit Rev Biochem Mol Biol. 2013 Jan-Feb;48(1):51-68. doi: 10.3109/10409238.2012.738643. Epub 2012 Nov 19. Crit Rev Biochem Mol Biol. 2013. PMID: 23163351 Free PMC article. Review.
-
Cotranscriptional RNA processing and modification in plants.Plant Cell. 2023 May 29;35(6):1654-1670. doi: 10.1093/plcell/koac309. Plant Cell. 2023. PMID: 36259932 Free PMC article. Review.
References
-
- Kim VN. MicroRNA biogenesis: coordinated cropping and dicing. Nat Rev Mol Cell Biol. 2005;6:376–385. - PubMed
-
- Denli AM, Tops BBJ, Plasterk RHA, Ketting RF, Hannon GJ. Processing of primary microRNAs by the microprocessor complex. Nature. 2004;432:231–235. - PubMed
-
- Gregory RI, et al. The Microprocessor complex mediates the genesis of microRNAs. Nature. 2004;432:235–240. - PubMed
-
- Han J, et al. Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex. Cell. 2006;125:887–901. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
