

A network of hydrophobic residues impeding helix alphaC rotation maintains latency of kinase Gcn2, which phosphorylates the alpha subunit of translation initiation factor 2
- PMID: 19114556
- PMCID: PMC2648240
- DOI: 10.1128/MCB.01446-08
A network of hydrophobic residues impeding helix alphaC rotation maintains latency of kinase Gcn2, which phosphorylates the alpha subunit of translation initiation factor 2
Abstract
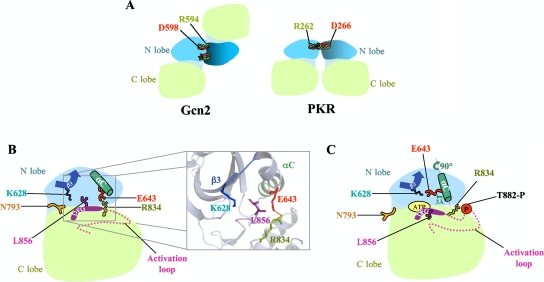
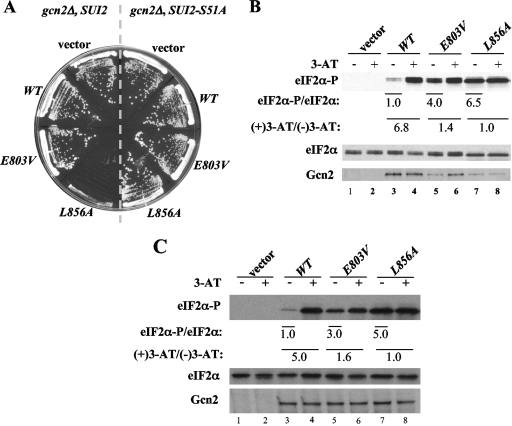
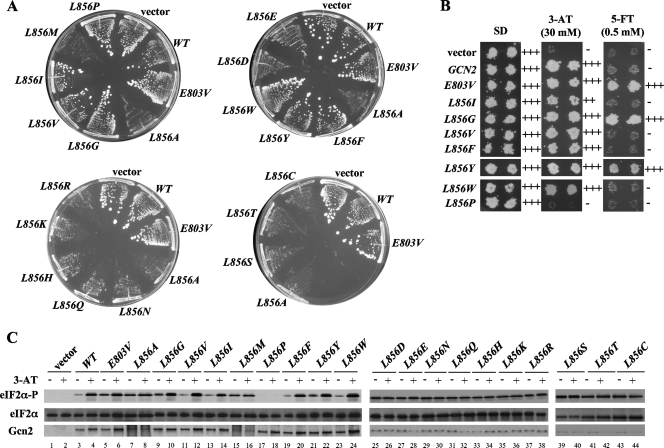
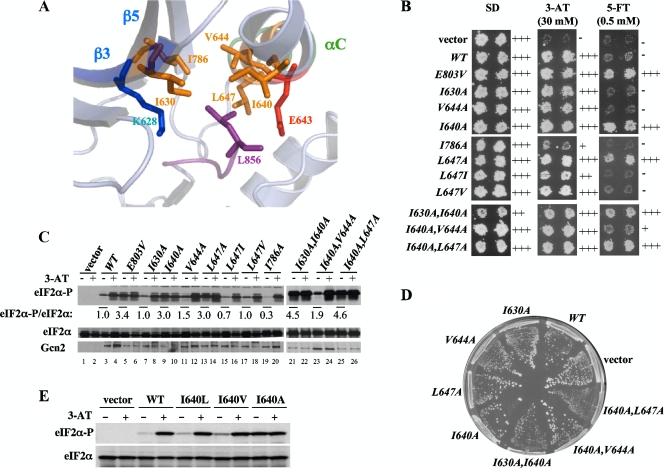
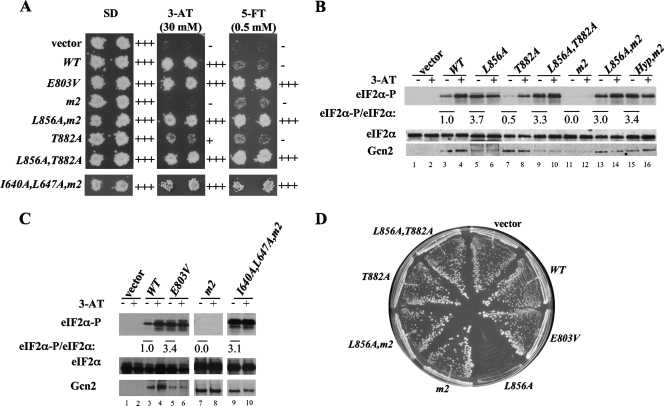
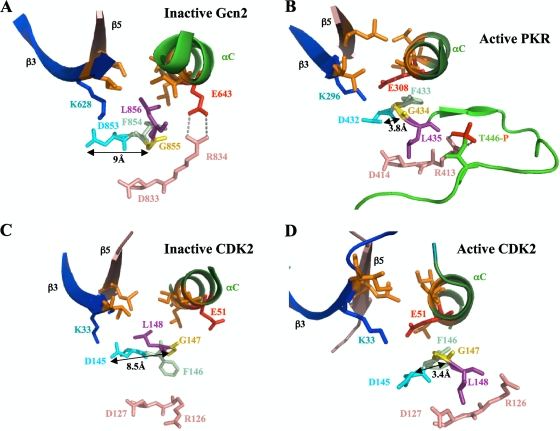
Kinase Gcn2 is activated by amino acid starvation and downregulates translation initiation by phosphorylating the alpha subunit of translation initiation factor 2 (eIF2alpha). The Gcn2 kinase domain (KD) is inert and must be activated by tRNA binding to the adjacent regulatory domain. Previous work indicated that Saccharomyces cerevisiae Gcn2 latency results from inflexibility of the hinge connecting the N and C lobes and a partially obstructed ATP-binding site in the KD. Here, we provide strong evidence that a network of hydrophobic interactions centered on Leu-856 also promotes latency by constraining helix alphaC rotation in the KD in a manner relieved during amino acid starvation by tRNA binding and autophosphorylation of Thr-882 in the activation loop. Thus, we show that mutationally disrupting the hydrophobic network in various ways constitutively activates eIF2alpha phosphorylation in vivo and bypasses the requirement for a key tRNA binding motif (m2) and Thr-882 in Gcn2. In particular, replacing Leu-856 with any nonhydrophobic residue activates Gcn2, while substitutions with various hydrophobic residues maintain kinase latency. We further provide strong evidence that parallel, back-to-back dimerization of the KD is a step on the Gcn2 activation pathway promoted by tRNA binding and autophosphorylation. Remarkably, mutations that disrupt the L856 hydrophobic network or enhance hinge flexibility eliminate the need for the conserved salt bridge at the parallel dimer interface, implying that KD dimerization facilitates the reorientation of alphaC and remodeling of the active site for enhanced ATP binding and catalysis. We propose that hinge remodeling, parallel dimerization, and reorientation of alphaC are mutually reinforcing conformational transitions stimulated by tRNA binding and secured by the ensuing autophosphorylation of T882 for stable kinase activation.
Figures



Similar articles
-
Evidence that eukaryotic translation elongation factor 1A (eEF1A) binds the Gcn2 protein C terminus and inhibits Gcn2 activity.J Biol Chem. 2011 Oct 21;286(42):36568-79. doi: 10.1074/jbc.M111.248898. Epub 2011 Aug 17. J Biol Chem. 2011. PMID: 21849502 Free PMC article.
-
Enhanced interaction between pseudokinase and kinase domains in Gcn2 stimulates eIF2α phosphorylation in starved cells.PLoS Genet. 2014 May 8;10(5):e1004326. doi: 10.1371/journal.pgen.1004326. eCollection 2014 May. PLoS Genet. 2014. PMID: 24811037 Free PMC article.
-
Interaction between the tRNA-binding and C-terminal domains of Yeast Gcn2 regulates kinase activity in vivo.PLoS Genet. 2015 Feb 19;11(2):e1004991. doi: 10.1371/journal.pgen.1004991. eCollection 2015 Feb. PLoS Genet. 2015. PMID: 25695491 Free PMC article.
-
New functions of protein kinase Gcn2 in yeast and mammals.IUBMB Life. 2012 Dec;64(12):971-4. doi: 10.1002/iub.1090. Epub 2012 Nov 5. IUBMB Life. 2012. PMID: 23129244 Review.
-
Regulation of translation initiation by amino acids in eukaryotic cells.Prog Mol Subcell Biol. 2001;26:155-84. doi: 10.1007/978-3-642-56688-2_6. Prog Mol Subcell Biol. 2001. PMID: 11575165 Review.
Cited by
-
Eukaryotic initiation factor 2 phosphorylation and translational control in metabolism.Adv Nutr. 2012 May 1;3(3):307-21. doi: 10.3945/an.112.002113. Adv Nutr. 2012. PMID: 22585904 Free PMC article. Review.
-
General control nonderepressible 2 (GCN2) as a therapeutic target in age-related diseases.Front Aging. 2024 Sep 10;5:1447370. doi: 10.3389/fragi.2024.1447370. eCollection 2024. Front Aging. 2024. PMID: 39319345 Free PMC article. Review.
-
Evidence that eukaryotic translation elongation factor 1A (eEF1A) binds the Gcn2 protein C terminus and inhibits Gcn2 activity.J Biol Chem. 2011 Oct 21;286(42):36568-79. doi: 10.1074/jbc.M111.248898. Epub 2011 Aug 17. J Biol Chem. 2011. PMID: 21849502 Free PMC article.
-
Integrated Stress Responses to Bacterial Pathogenesis Patterns.Front Immunol. 2018 Jun 7;9:1306. doi: 10.3389/fimmu.2018.01306. eCollection 2018. Front Immunol. 2018. PMID: 29930559 Free PMC article. Review.
-
Enhanced interaction between pseudokinase and kinase domains in Gcn2 stimulates eIF2α phosphorylation in starved cells.PLoS Genet. 2014 May 8;10(5):e1004326. doi: 10.1371/journal.pgen.1004326. eCollection 2014 May. PLoS Genet. 2014. PMID: 24811037 Free PMC article.
References
-
- Adams, J., P. Huang, and D. Patrick. 2002. A strategy for the design of multiplex inhibitors for kinase-mediated signaling in angiogenesis. Curr. Opin. Chem. Biol. 6486-492. - PubMed
-
- Brown, N. R., M. E. Noble, J. A. Endicott, and L. N. Johnson. 1999. The structural basis for specificity of substrate and recruitment peptides for cyclin-dependent kinases. Nat. Cell Biol. 1438-443. - PubMed
-
- Cai, R., and B. R. Williams. 1998. Mutations in the double-stranded RNA-activated protein kinase insert region that uncouple catalysis from eIF2alpha binding. J. Biol. Chem. 27311274-11280. - PubMed
-
- Costa-Mattioli, M., D. Gobert, H. Harding, B. Herdy, M. Azzi, M. Bruno, M. Bidinosti, C. Ben Mamou, E. Marcinkiewicz, M. Yoshida, H. Imataka, A. C. Cuello, N. Seidah, W. Sossin, J. C. Lacaille, D. Ron, K. Nader, and N. Sonenberg. 2005. Translational control of hippocampal synaptic plasticity and memory by the eIF2alpha kinase GCN2. Nature 4361166-1173. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases