

Nutrient-regulated antisense and intragenic RNAs modulate a signal transduction pathway in yeast
- PMID: 19108609
- PMCID: PMC2605928
- DOI: 10.1371/journal.pbio.0060326
Nutrient-regulated antisense and intragenic RNAs modulate a signal transduction pathway in yeast
Abstract
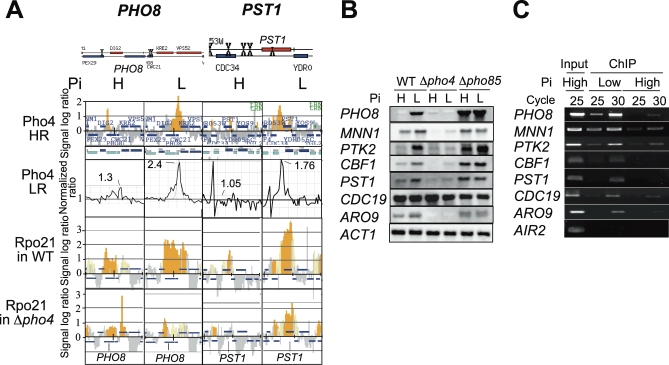
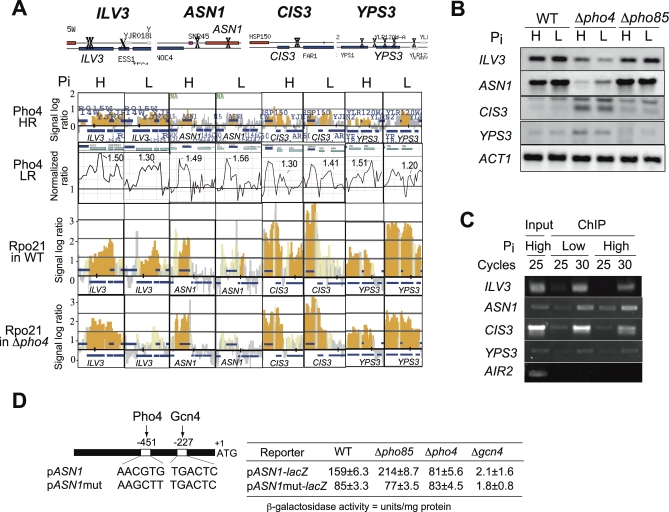
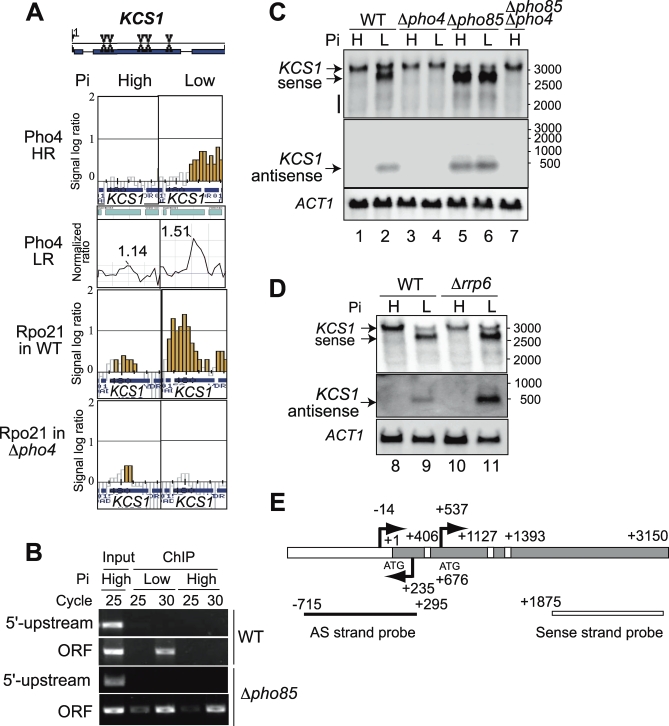
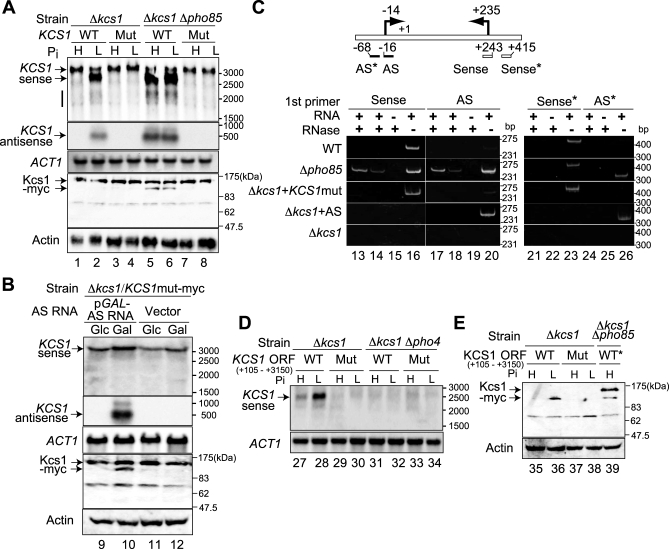
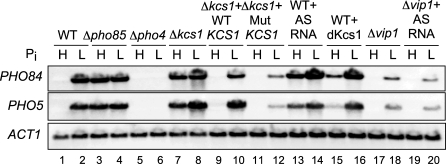
The budding yeast Saccharomyces cerevisiae alters its gene expression profile in response to a change in nutrient availability. The PHO system is a well-studied case in the transcriptional regulation responding to nutritional changes in which a set of genes (PHO genes) is expressed to activate inorganic phosphate (Pi) metabolism for adaptation to Pi starvation. Pi starvation triggers an inhibition of Pho85 kinase, leading to migration of unphosphorylated Pho4 transcriptional activator into the nucleus and enabling expression of PHO genes. When Pi is sufficient, the Pho85 kinase phosphorylates Pho4, thereby excluding it from the nucleus and resulting in repression (i.e., lack of transcription) of PHO genes. The Pho85 kinase has a role in various cellular functions other than regulation of the PHO system in that Pho85 monitors whether environmental conditions are adequate for cell growth and represses inadequate (untimely) responses in these cellular processes. In contrast, Pho4 appears to activate some genes involved in stress response and is required for G1 arrest caused by DNA damage. These facts suggest the antagonistic function of these two players on a more general scale when yeast cells must cope with stress conditions. To explore general involvement of Pho4 in stress response, we tried to identify Pho4-dependent genes by a genome-wide mapping of Pho4 and Rpo21 binding (Rpo21 being the largest subunit of RNA polymerase II) using a yeast tiling array. In the course of this study, we found Pi- and Pho4-regulated intragenic and antisense RNAs that could modulate the Pi signal transduction pathway. Low-Pi signal is transmitted via certain inositol polyphosphate (IP) species (IP7) that are synthesized by Vip1 IP6 kinase. We have shown that Pho4 activates the transcription of antisense and intragenic RNAs in the KCS1 locus to down-regulate the Kcs1 activity, another IP6 kinase, by producing truncated Kcs1 protein via hybrid formation with the KCS1 mRNA and translation of the intragenic RNA, thereby enabling Vip1 to utilize more IP6 to synthesize IP7 functioning in low-Pi signaling. Because Kcs1 also can phosphorylate these IP7 species to synthesize IP8, reduction in Kcs1 activity can ensure accumulation of the IP7 species, leading to further stimulation of low-Pi signaling (i.e., forming a positive feedback loop). We also report that genes apparently not involved in the PHO system are regulated by Pho4 either dependent upon or independent of the Pi conditions, and many of the latter genes are involved in stress response. In S. cerevisiae, a large-scale cDNA analysis and mapping of RNA polymerase II binding using a high-resolution tiling array have identified a large number of antisense RNA species whose functions are yet to be clarified. Here we have shown that nutrient-regulated antisense and intragenic RNAs as well as direct regulation of structural gene transcription function in the response to nutrient availability. Our findings also imply that Pho4 is present in the nucleus even under high-Pi conditions to activate or repress transcription, which challenges our current understanding of Pho4 regulation.
Conflict of interest statement
Figures
Similar articles
-
A systematic genetic screen for genes involved in sensing inorganic phosphate availability in Saccharomyces cerevisiae.PLoS One. 2017 May 17;12(5):e0176085. doi: 10.1371/journal.pone.0176085. eCollection 2017. PLoS One. 2017. PMID: 28520786 Free PMC article.
-
Evolution of reduced co-activator dependence led to target expansion of a starvation response pathway.Elife. 2017 May 9;6:e25157. doi: 10.7554/eLife.25157. Elife. 2017. PMID: 28485712 Free PMC article.
-
Transcriptional repression by the Pho4 transcription factor controls the timing of SNZ1 expression.Eukaryot Cell. 2008 Jun;7(6):949-57. doi: 10.1128/EC.00366-07. Epub 2008 Apr 11. Eukaryot Cell. 2008. PMID: 18408055 Free PMC article.
-
Kcs1 and Vip1: The Key Enzymes behind Inositol Pyrophosphate Signaling in Saccharomyces cerevisiae.Biomolecules. 2024 Jan 26;14(2):152. doi: 10.3390/biom14020152. Biomolecules. 2024. PMID: 38397389 Free PMC article. Review.
-
The response to heat shock and oxidative stress in Saccharomyces cerevisiae.Genetics. 2012 Apr;190(4):1157-95. doi: 10.1534/genetics.111.128033. Epub 2011 Dec 29. Genetics. 2012. PMID: 22209905 Free PMC article. Review.
Cited by
-
Phosphate-activated cyclin-dependent kinase stabilizes G1 cyclin to trigger cell cycle entry.Mol Cell Biol. 2013 Apr;33(7):1273-84. doi: 10.1128/MCB.01556-12. Epub 2013 Jan 22. Mol Cell Biol. 2013. PMID: 23339867 Free PMC article.
-
Strand-specific RNA sequencing reveals extensive regulated long antisense transcripts that are conserved across yeast species.Genome Biol. 2010;11(8):R87. doi: 10.1186/gb-2010-11-8-r87. Epub 2010 Aug 26. Genome Biol. 2010. PMID: 20796282 Free PMC article.
-
Genome-Wide Analysis of Nutrient Signaling Pathways Conserved in Arbuscular Mycorrhizal Fungi.Microorganisms. 2021 Jul 22;9(8):1557. doi: 10.3390/microorganisms9081557. Microorganisms. 2021. PMID: 34442636 Free PMC article.
-
Differential impact of nutrition on developmental and metabolic gene expression during fruiting body development in Neurospora crassa.Fungal Genet Biol. 2012 May;49(5):405-13. doi: 10.1016/j.fgb.2012.03.004. Epub 2012 Mar 26. Fungal Genet Biol. 2012. PMID: 22469835 Free PMC article.
-
The NPR/Hal family of protein kinases in yeasts: biological role, phylogeny and regulation under environmental challenges.Comput Struct Biotechnol J. 2022 Oct 15;20:5698-5712. doi: 10.1016/j.csbj.2022.10.006. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36320937 Free PMC article. Review.
References
-
- Carroll AS, O'Shea EK. Pho85 and signaling environmental conditions. Trends Biochem Sci. 2002;27:87–93. - PubMed
-
- Wilson WA, Roach PJ. Nutrient-regulated protein kinases in budding yeast. Cell. 2002;111:155–158. - PubMed
-
- Oshima Y. The phosphate system in Saccharomyces cerevisiae . Genes Genet Syst. 1997;72:323–334. - PubMed
-
- Kaffman A, Rank NM, O'Neill EM, Huang LS, O'Shea EK. The receptor Msn5 exports the phosphorylated transcription factor Pho4 out of the nucleus. Nature. 1998b;396:482–486. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
