

Migratory and lymphoid-resident dendritic cells cooperate to efficiently prime naive CD4 T cells
- PMID: 18951047
- PMCID: PMC2746692
- DOI: 10.1016/j.immuni.2008.08.013
Migratory and lymphoid-resident dendritic cells cooperate to efficiently prime naive CD4 T cells
Abstract
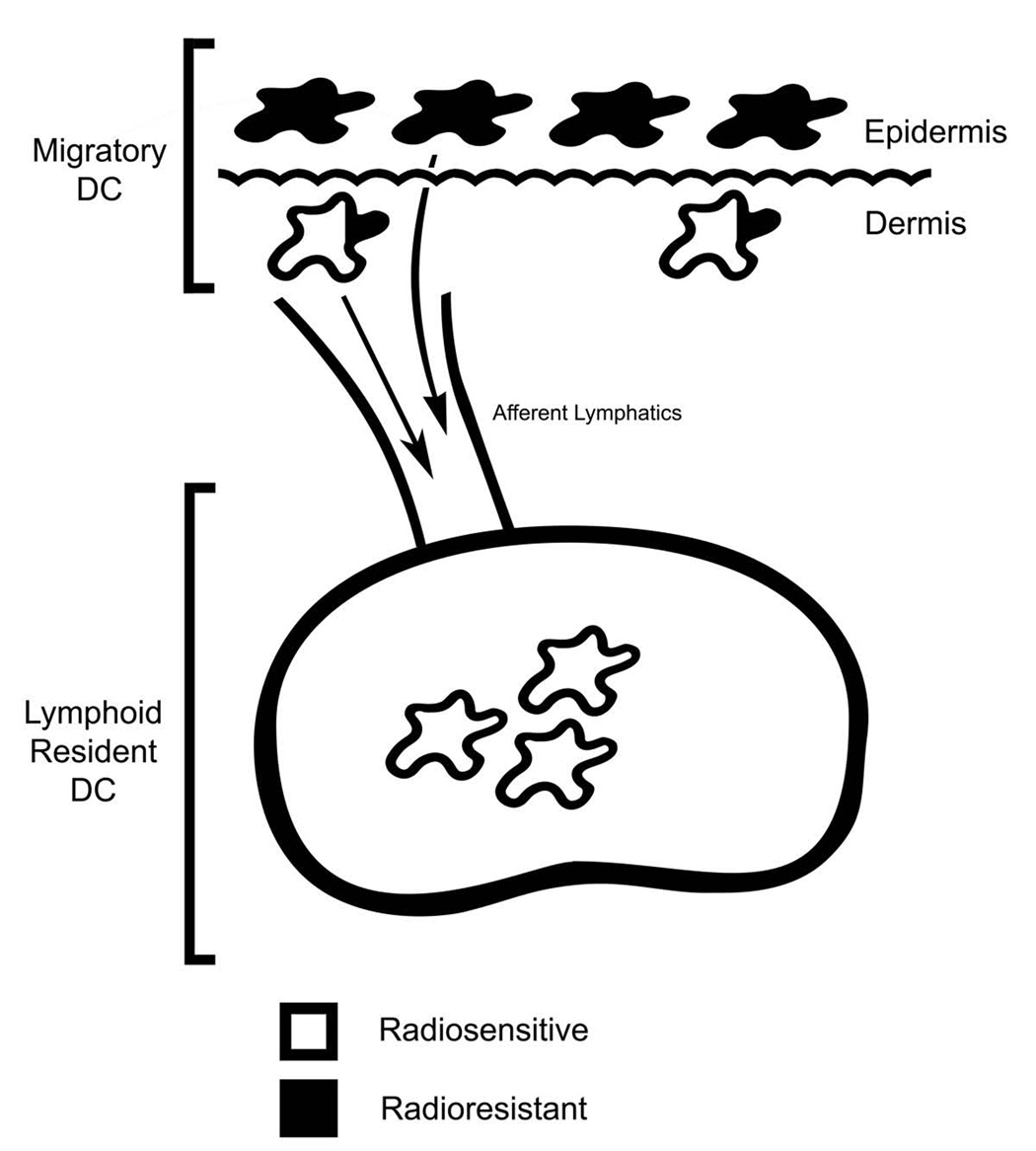
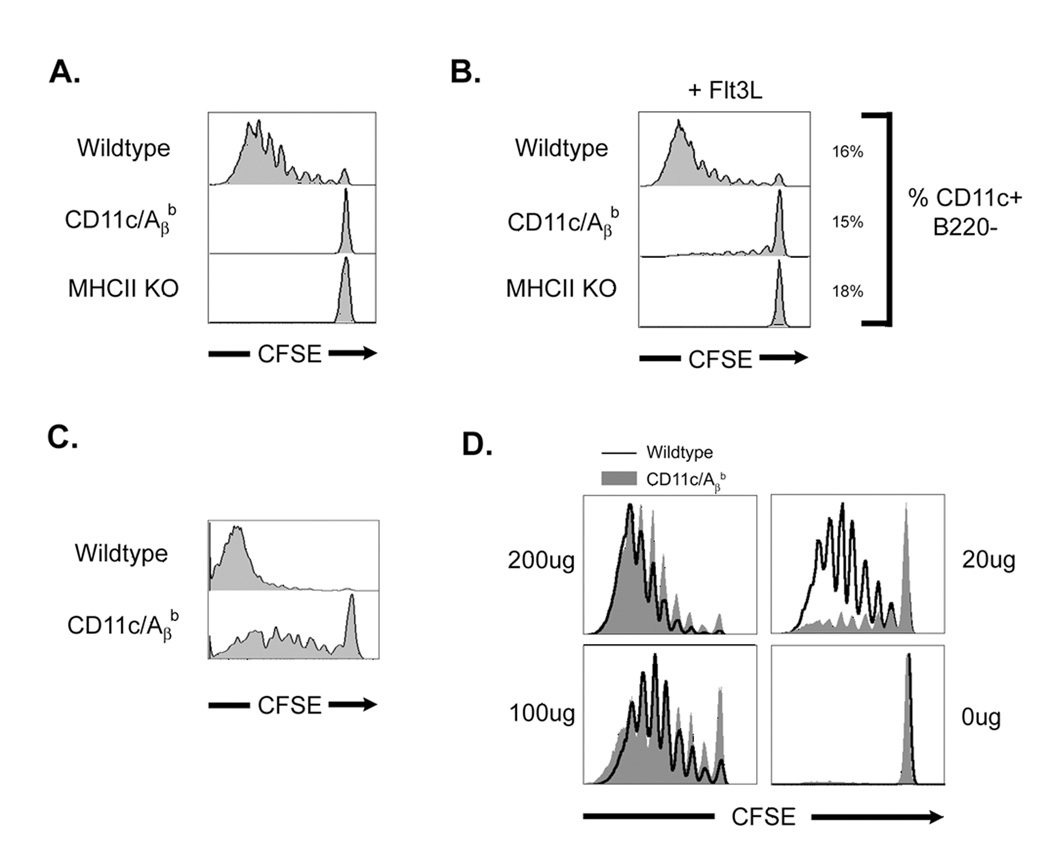
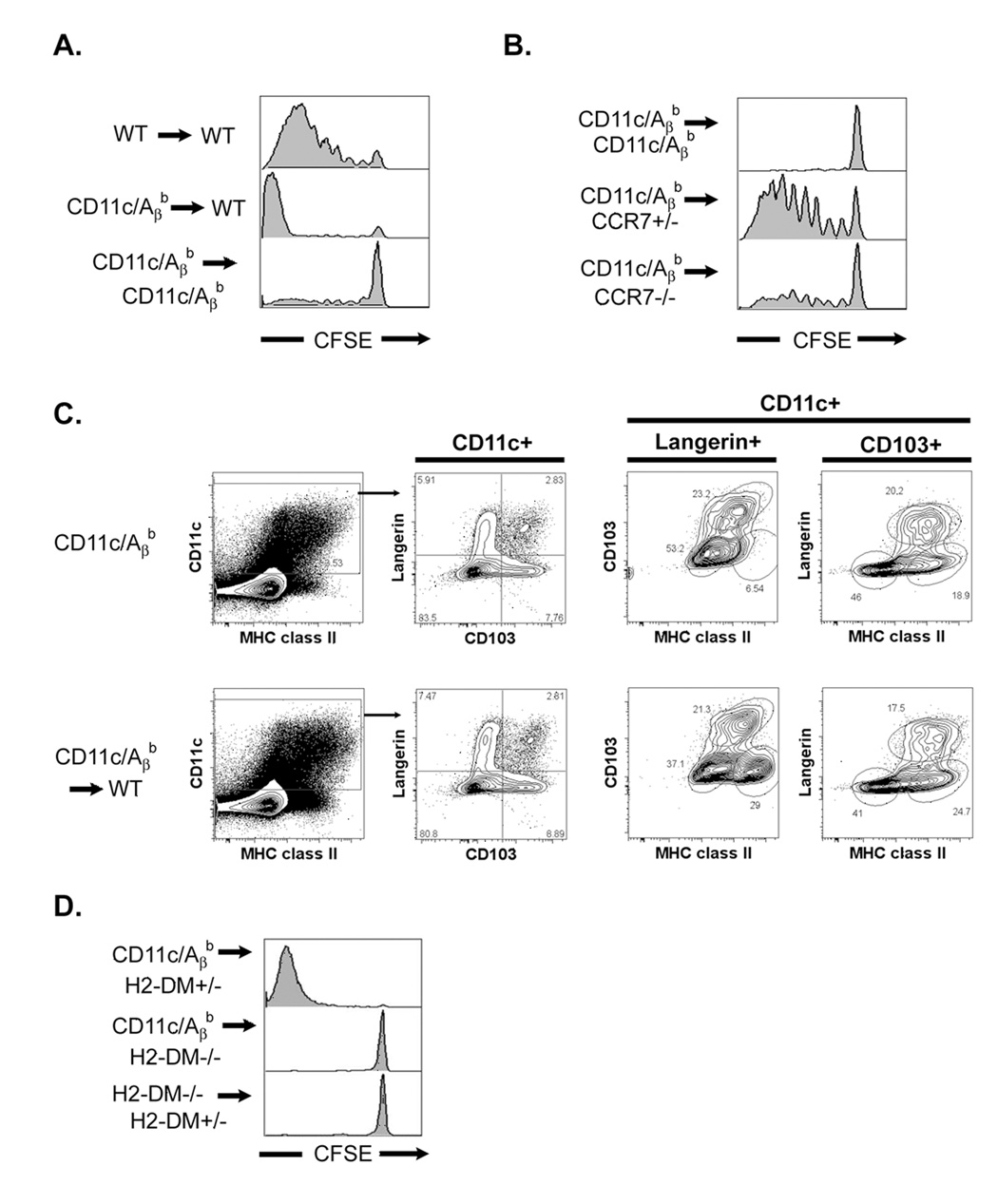
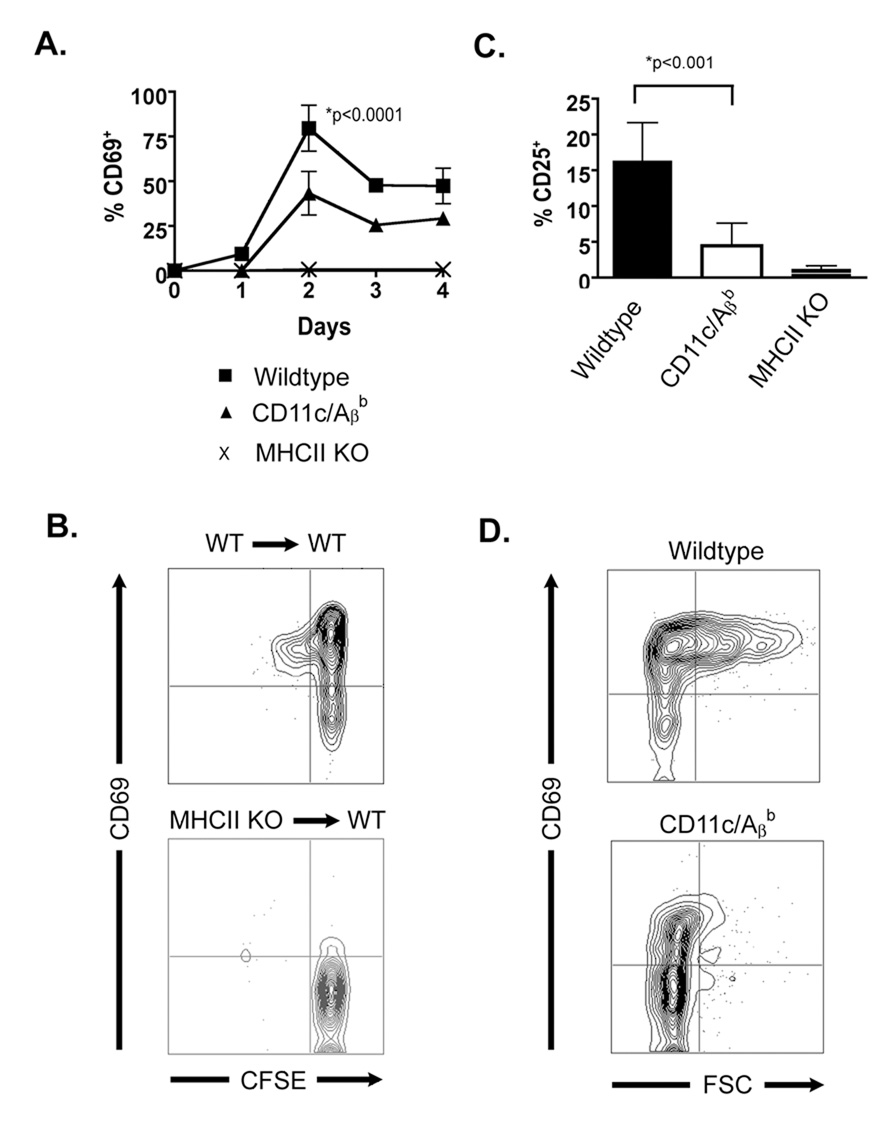
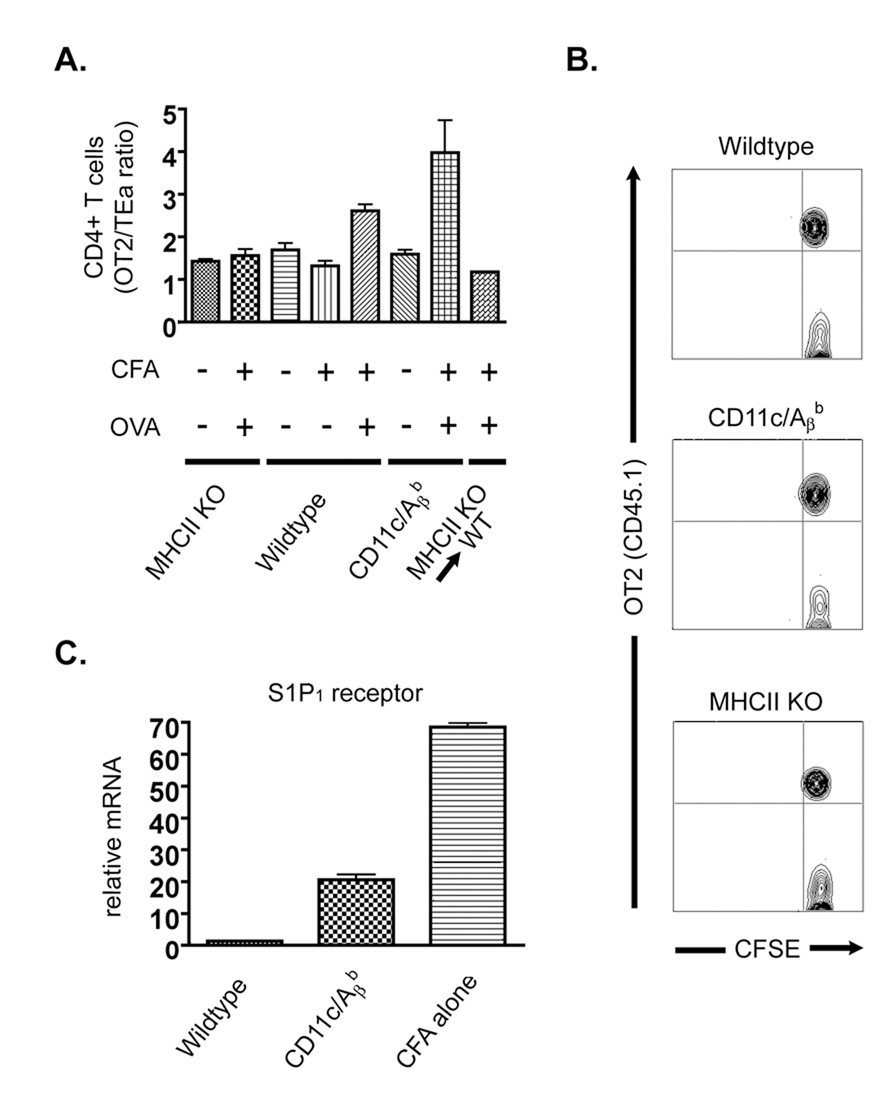
To initiate an adaptive immune response, rare antigen-specific naive CD4(+) T cells must interact with equally rare dendritic cells (DCs) bearing cognate peptide-major histocompatibility complex (MHC) complexes. Lymph nodes (LNs) draining the site of antigen entry are populated by lymphoid-resident DCs as well as DCs that have immigrated from tissues, although the requirement for each population in initiating the T cell response remains unclear. Here, we show that antigen processing and presentation by both lymphoid-resident and migratory DCs was required for clonal selection and expansion of CD4(+) T cells after subcutaneous immunization. Early antigen presentation by lymphoid-resident DCs initiated activation and trapping of antigen-specific T cells in the draining LN, without sufficing for clonal expansion. Migratory DCs, however, interacted with the CD4(+) T cells retained in the LN to induce proliferation. Therefore, distinct DC subsets cooperate to alert and trap the appropriate cell and then license its expansion and differentiation.
Figures


Similar articles
-
Splenic dendritic cells pulsed with Ixodes ricinus tick saliva prime naive CD4+T to induce Th2 cell differentiation in vitro and in vivo.Int Immunol. 2007 Apr;19(4):535-43. doi: 10.1093/intimm/dxm019. Epub 2007 Mar 6. Int Immunol. 2007. PMID: 17344202
-
Spatiotemporally Distinct Interactions with Dendritic Cell Subsets Facilitates CD4+ and CD8+ T Cell Activation to Localized Viral Infection.Immunity. 2015 Sep 15;43(3):554-65. doi: 10.1016/j.immuni.2015.07.020. Epub 2015 Aug 18. Immunity. 2015. PMID: 26297566
-
Antigen presentation by dendritic cells in renal lymph nodes is linked to systemic and local injury to the kidney.Kidney Int. 2005 Sep;68(3):1096-108. doi: 10.1111/j.1523-1755.2005.00502.x. Kidney Int. 2005. PMID: 16105040
-
Studying interactions between dendritic cells and T cells in vivo.Curr Opin Immunol. 2019 Jun;58:24-30. doi: 10.1016/j.coi.2019.02.002. Epub 2019 Mar 15. Curr Opin Immunol. 2019. PMID: 30884422 Free PMC article. Review.
-
Compartmentalization of dendritic cell and T-cell interactions in the lymph node: Anatomy of T-cell fate decisions.Immunol Rev. 2019 May;289(1):84-100. doi: 10.1111/imr.12758. Immunol Rev. 2019. PMID: 30977197 Free PMC article. Review.
Cited by
-
Multistage T cell-dendritic cell interactions control optimal CD4 T cell activation through the ADAP-SKAP55-signaling module.J Immunol. 2013 Sep 1;191(5):2372-83. doi: 10.4049/jimmunol.1300107. Epub 2013 Aug 5. J Immunol. 2013. PMID: 23918975 Free PMC article.
-
Cyclophosphamide enhances immunity by modulating the balance of dendritic cell subsets in lymphoid organs.Blood. 2010 Jun 3;115(22):4384-92. doi: 10.1182/blood-2009-11-251231. Epub 2010 Feb 12. Blood. 2010. PMID: 20154220 Free PMC article.
-
CD8α dendritic cells drive establishment of HSV-1 latency.PLoS One. 2014 Apr 2;9(4):e93444. doi: 10.1371/journal.pone.0093444. eCollection 2014. PLoS One. 2014. PMID: 24695322 Free PMC article.
-
Immunomodulatory Properties of Amniotic Membrane Derivatives and Their Potential in Regenerative Medicine.Curr Diab Rep. 2020 Jun 10;20(8):31. doi: 10.1007/s11892-020-01316-w. Curr Diab Rep. 2020. PMID: 32519069 Free PMC article. Review.
-
Lymphatic function and immune regulation in health and disease.Lymphat Res Biol. 2013 Sep;11(3):136-43. doi: 10.1089/lrb.2013.0012. Epub 2013 Sep 11. Lymphat Res Biol. 2013. PMID: 24024577 Free PMC article. Review. No abstract available.
References
-
- Allan RS, Smith CM, Belz GT, van Lint AL, Wakim LM, Heath WR, Carbone FR. Epidermal viral immunity induced by CD8alpha+ dendritic cells but not by Langerhans cells. Science. 2003;301:1925–1928. - PubMed
-
- Allan RS, Waithman J, Bedoui S, Jones CM, Villadangos JA, Zhan Y, Lew AM, Shortman K, Heath WR, Carbone FR. Migratory dendritic cells transfer antigen to a lymph node-resident dendritic cell population for efficient CTL priming. Immunity. 2006;25:153–162. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
