

53BP1 facilitates long-range DNA end-joining during V(D)J recombination
- PMID: 18931658
- PMCID: PMC3596817
- DOI: 10.1038/nature07476
53BP1 facilitates long-range DNA end-joining during V(D)J recombination
Abstract
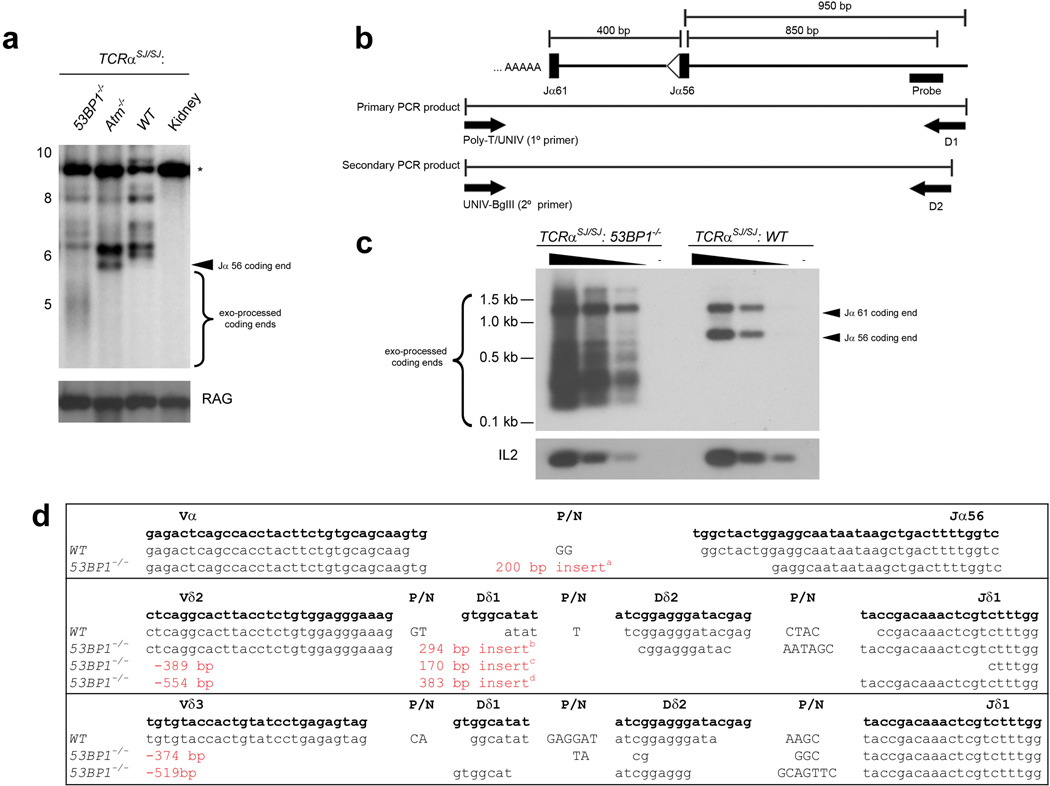
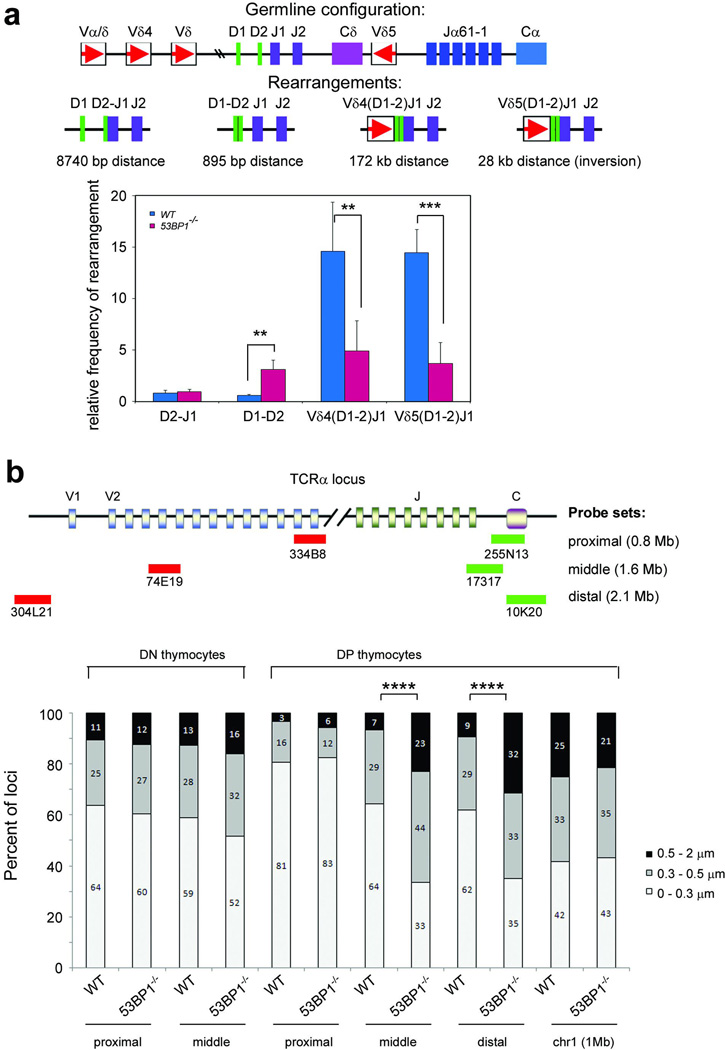
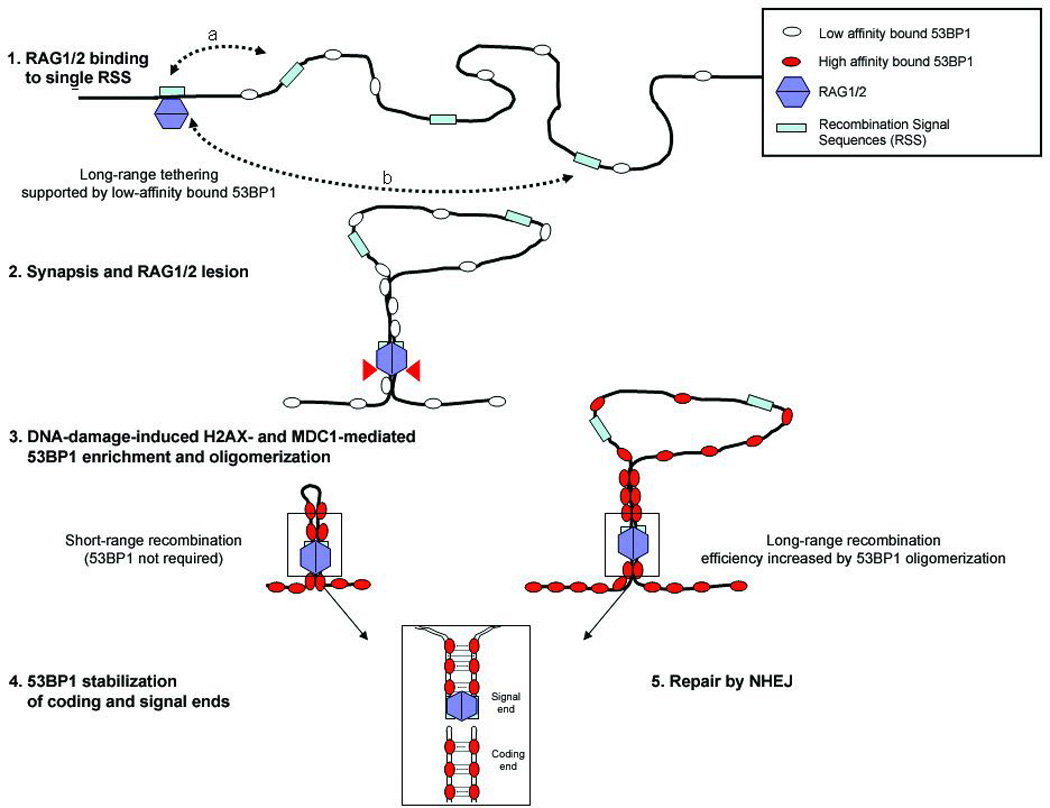
Variable, diversity and joining (V(D)J) recombination and class-switch recombination use overlapping but distinct non-homologous end joining pathways to repair DNA double-strand-break intermediates. 53BP1 is a DNA-damage-response protein that is rapidly recruited to sites of chromosomal double-strand breaks, where it seems to function in a subset of ataxia telangiectasia mutated (ATM) kinase-, H2A histone family member X (H2AX, also known as H2AFX)- and mediator of DNA damage checkpoint 1 (MDC1)-dependent events. A 53BP1-dependent end-joining pathway has been described that is dispensable for V(D)J recombination but essential for class-switch recombination. Here we report a previously unrecognized defect in the joining phase of V(D)J recombination in 53BP1-deficient lymphocytes that is distinct from that found in classical non-homologous-end-joining-, H2ax-, Mdc1- and Atm-deficient mice. Absence of 53BP1 leads to impairment of distal V-DJ joining with extensive degradation of unrepaired coding ends and episomal signal joint reintegration at V(D)J junctions. This results in apoptosis, loss of T-cell receptor alpha locus integrity and lymphopenia. Further impairment of the apoptotic checkpoint causes propagation of lymphocytes that have antigen receptor breaks. These data suggest a more general role for 53BP1 in maintaining genomic stability during long-range joining of DNA breaks.
Figures

Similar articles
-
Overlapping functions between XLF repair protein and 53BP1 DNA damage response factor in end joining and lymphocyte development.Proc Natl Acad Sci U S A. 2012 Mar 6;109(10):3903-8. doi: 10.1073/pnas.1120160109. Epub 2012 Feb 21. Proc Natl Acad Sci U S A. 2012. PMID: 22355127 Free PMC article.
-
53BP1 promotes non-homologous end joining of telomeres by increasing chromatin mobility.Nature. 2008 Nov 27;456(7221):524-8. doi: 10.1038/nature07433. Epub 2008 Oct 19. Nature. 2008. PMID: 18931659 Free PMC article.
-
ATM damage response and XLF repair factor are functionally redundant in joining DNA breaks.Nature. 2011 Jan 13;469(7329):250-4. doi: 10.1038/nature09604. Epub 2010 Dec 15. Nature. 2011. PMID: 21160472 Free PMC article.
-
53BP1-mediated DNA double strand break repair: insert bad pun here.DNA Repair (Amst). 2011 Oct 10;10(10):1071-6. doi: 10.1016/j.dnarep.2011.07.012. Epub 2011 Aug 24. DNA Repair (Amst). 2011. PMID: 21868291 Review.
-
Double-strand break repair: 53BP1 comes into focus.Nat Rev Mol Cell Biol. 2014 Jan;15(1):7-18. doi: 10.1038/nrm3719. Epub 2013 Dec 11. Nat Rev Mol Cell Biol. 2014. PMID: 24326623 Review.
Cited by
-
53BP1 mediates productive and mutagenic DNA repair through distinct phosphoprotein interactions.Cell. 2013 Jun 6;153(6):1266-80. doi: 10.1016/j.cell.2013.05.023. Epub 2013 May 30. Cell. 2013. PMID: 23727112 Free PMC article.
-
A two-step mechanism for TRF2-mediated chromosome-end protection.Nature. 2013 Feb 28;494(7438):502-5. doi: 10.1038/nature11873. Epub 2013 Feb 6. Nature. 2013. PMID: 23389450 Free PMC article.
-
Class switching and meiotic defects in mice lacking the E3 ubiquitin ligase RNF8.J Exp Med. 2010 May 10;207(5):973-81. doi: 10.1084/jem.20092308. Epub 2010 Apr 12. J Exp Med. 2010. PMID: 20385748 Free PMC article.
-
Rnf8 deficiency impairs class switch recombination, spermatogenesis, and genomic integrity and predisposes for cancer.J Exp Med. 2010 May 10;207(5):983-97. doi: 10.1084/jem.20092437. Epub 2010 Apr 12. J Exp Med. 2010. PMID: 20385750 Free PMC article.
-
Congenital bone marrow failure in DNA-PKcs mutant mice associated with deficiencies in DNA repair.J Cell Biol. 2011 Apr 18;193(2):295-305. doi: 10.1083/jcb.201009074. Epub 2011 Apr 11. J Cell Biol. 2011. PMID: 21482716 Free PMC article.
References
-
- Stucki M, Jackson SP. gammaH2AX and MDC1: anchoring the DNA-damage-response machinery to broken chromosomes. DNA repair. 2006;5:534–543. - PubMed
-
- Fernandez-Capetillo O, Lee A, Nussenzweig M, Nussenzweig A. H2AX: the histone guardian of the genome. DNA repair. 2004;3:959–967. - PubMed
-
- Manis JP, et al. 53BP1 links DNA damage-response pathways to immunoglobulin heavy chain class-switch recombination. Nature immunology. 2004;5:481–487. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
