

Structure of a complex of the ATPase SecA and the protein-translocation channel
- PMID: 18923516
- PMCID: PMC7164768
- DOI: 10.1038/nature07335
Structure of a complex of the ATPase SecA and the protein-translocation channel
Abstract
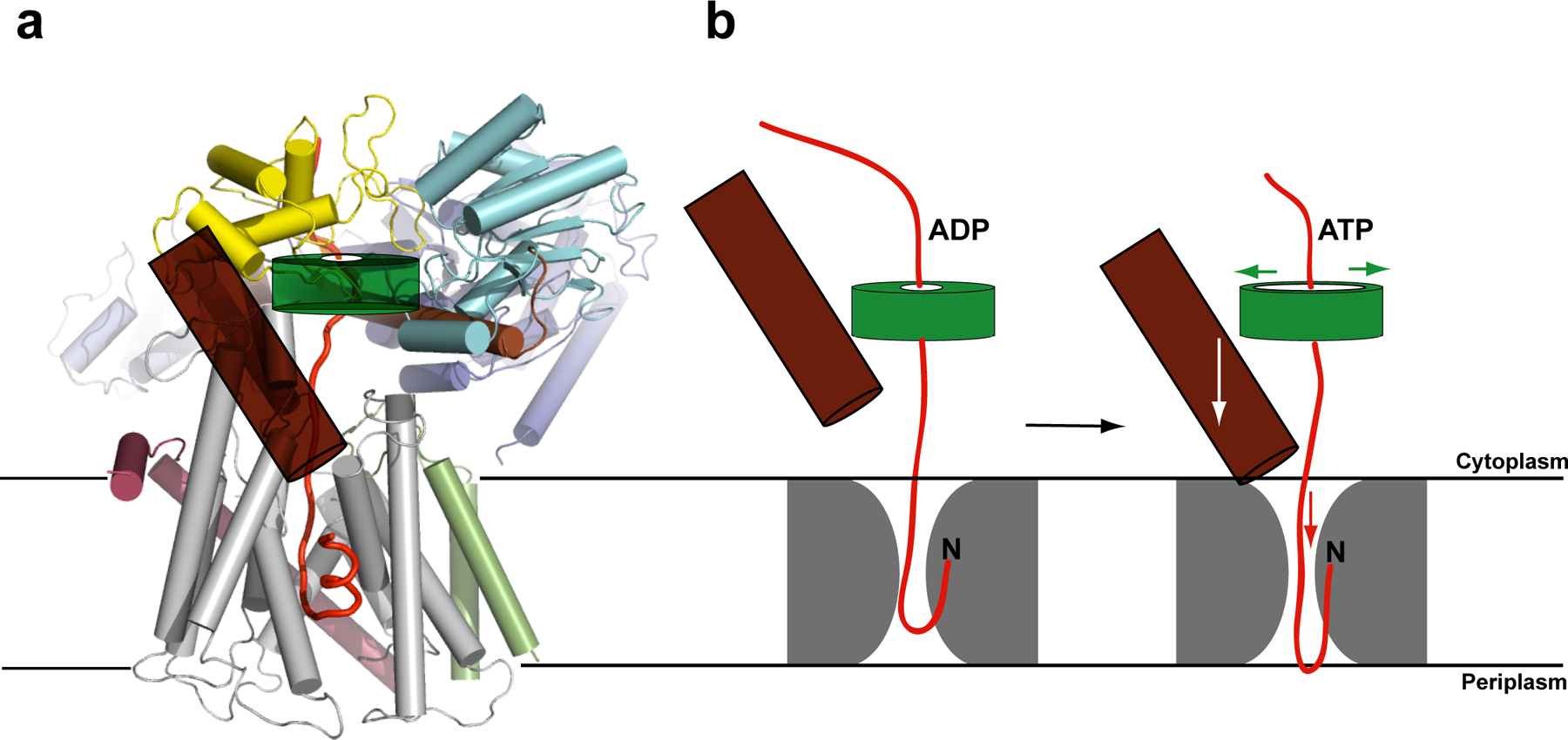
Most proteins are secreted from bacteria by the interaction of the cytoplasmic SecA ATPase with a membrane channel, formed by the heterotrimeric SecY complex. Here we report the crystal structure of SecA bound to the SecY complex, with a maximum resolution of 4.5 ångström (A), obtained for components from Thermotoga maritima. One copy of SecA in an intermediate state of ATP hydrolysis is bound to one molecule of the SecY complex. Both partners undergo important conformational changes on interaction. The polypeptide-cross-linking domain of SecA makes a large conformational change that could capture the translocation substrate in a 'clamp'. Polypeptide movement through the SecY channel could be achieved by the motion of a 'two-helix finger' of SecA inside the cytoplasmic funnel of SecY, and by the coordinated tightening and widening of SecA's clamp above the SecY pore. SecA binding generates a 'window' at the lateral gate of the SecY channel and it displaces the plug domain, preparing the channel for signal sequence binding and channel opening.
Figures
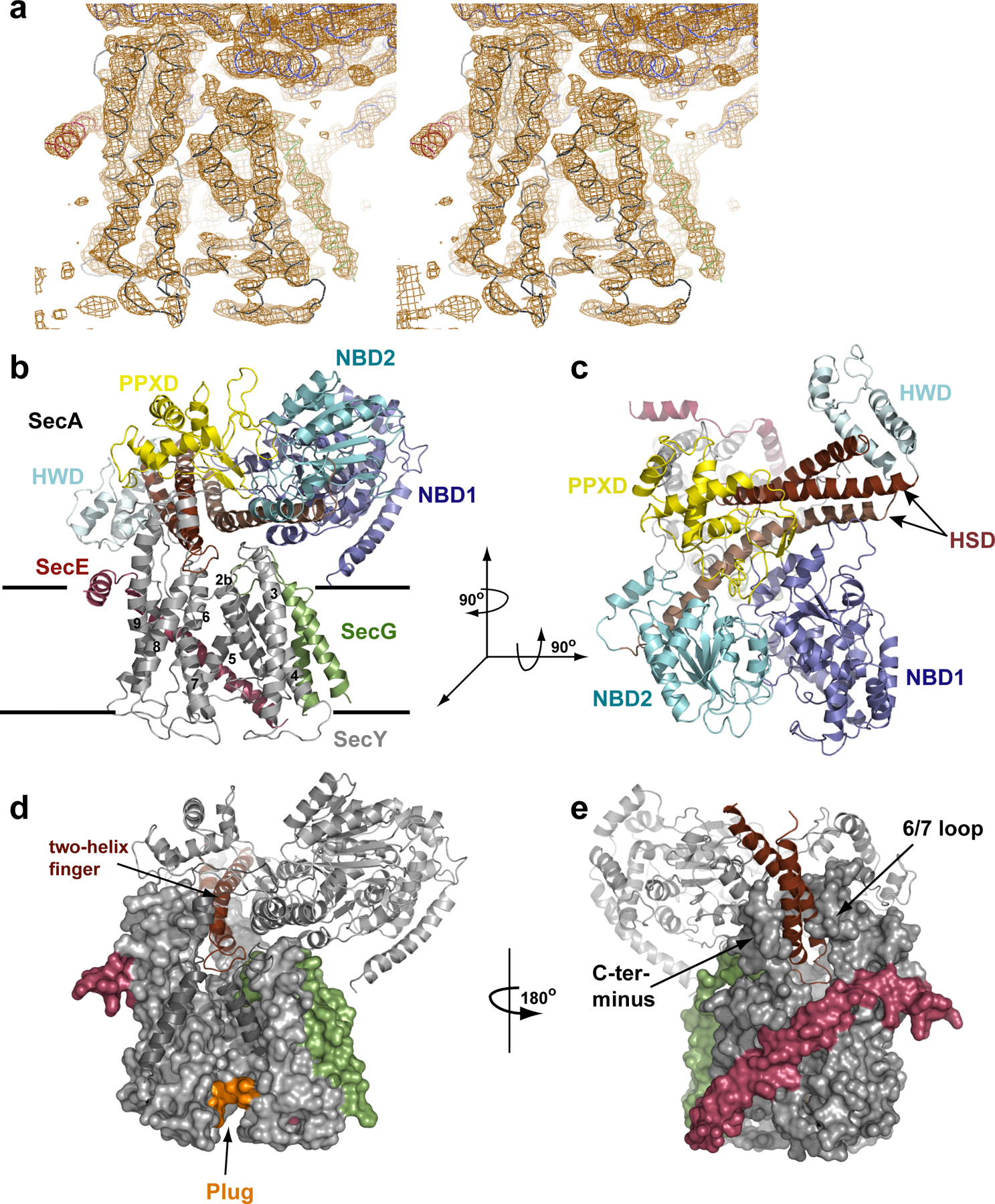
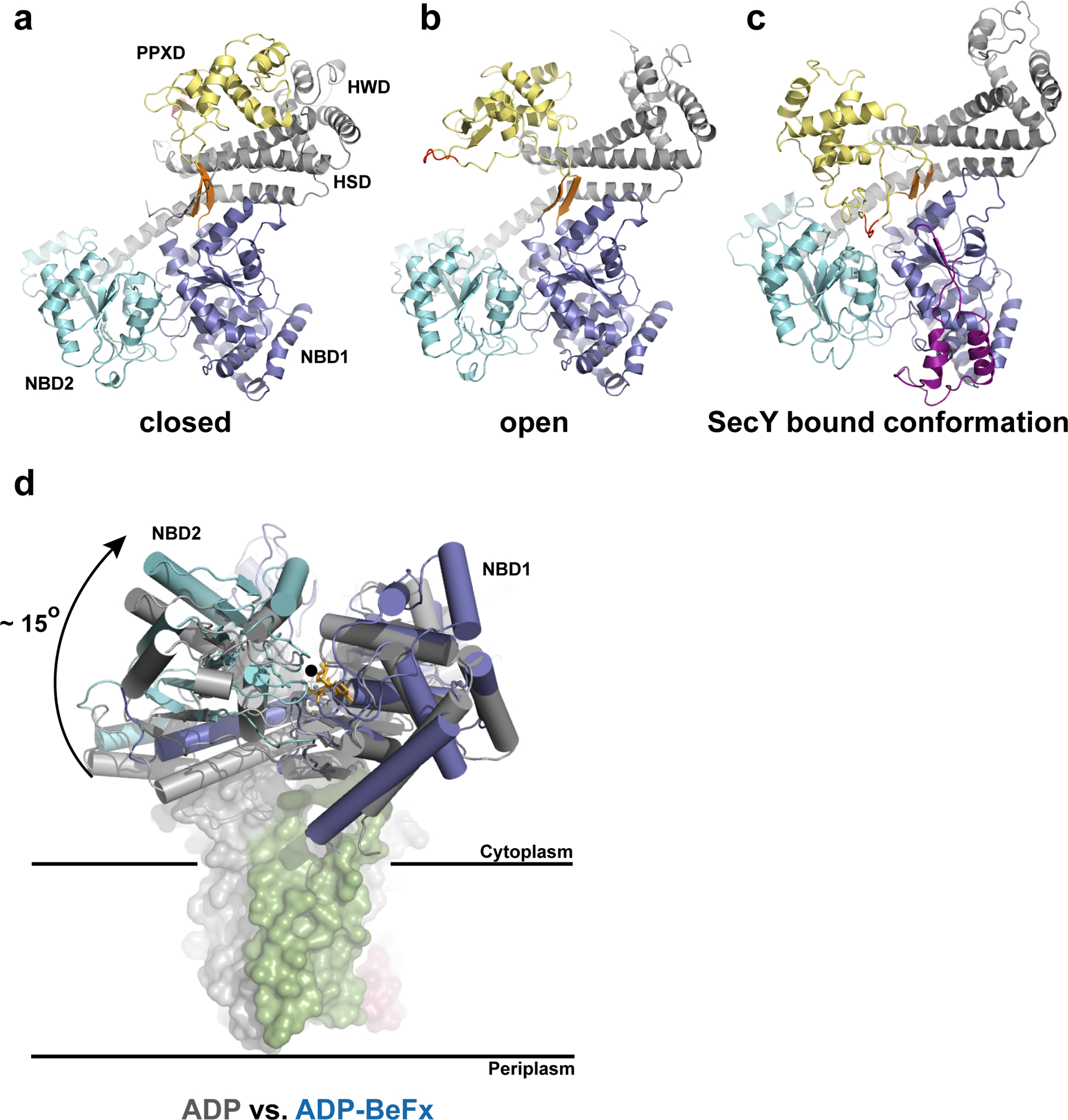
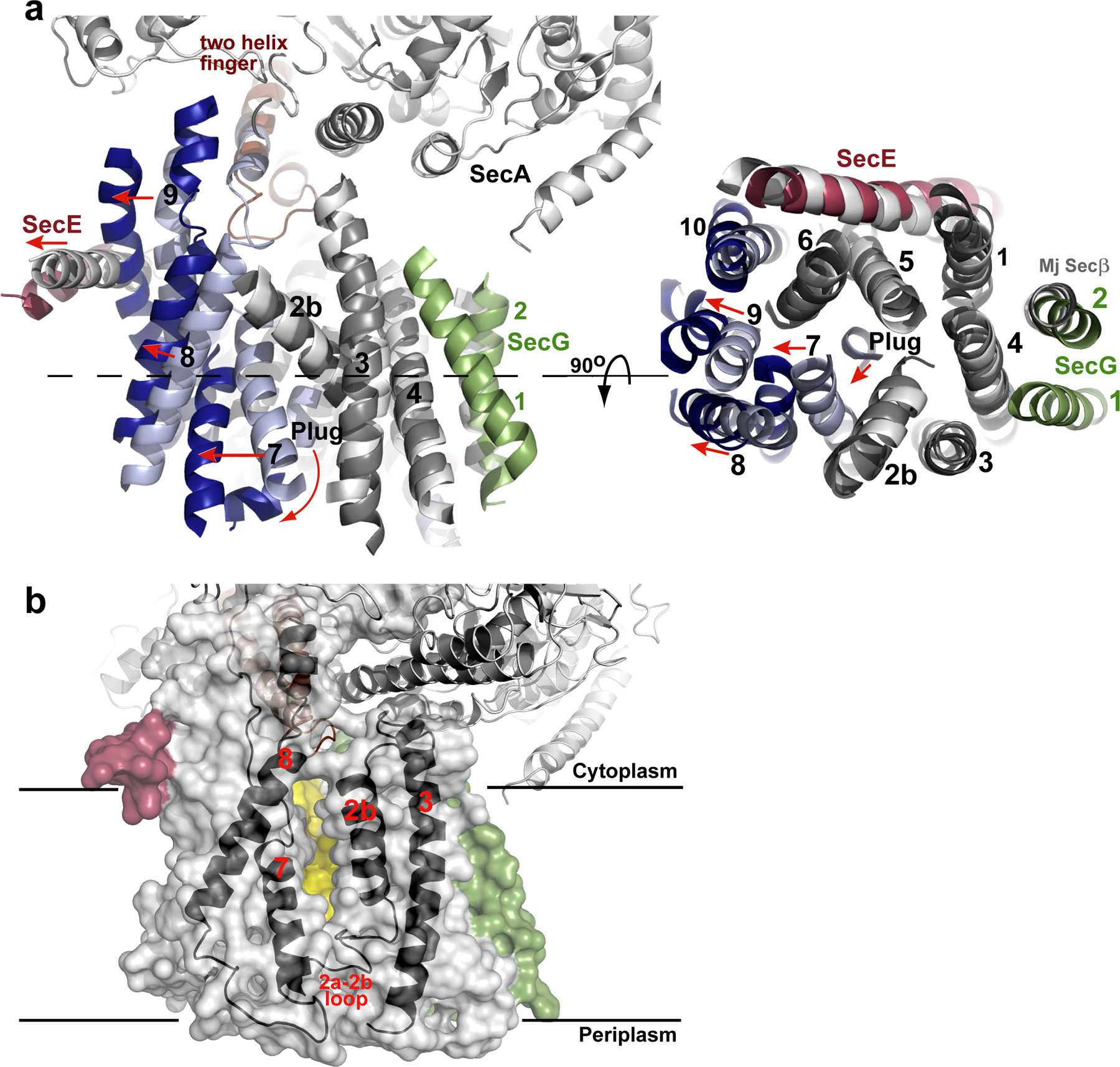
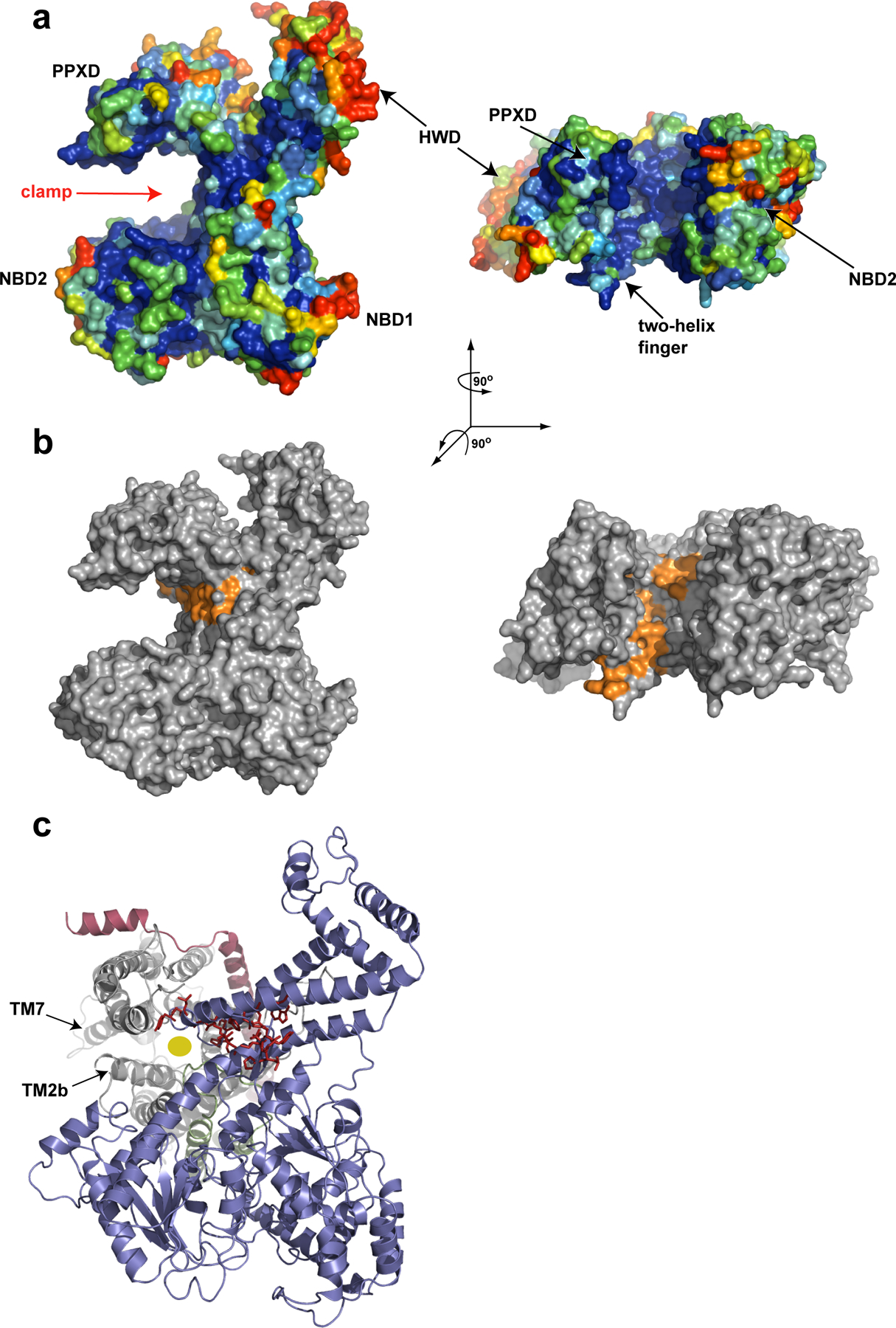
Comment in
-
Structural biology: Clamour for a kiss.Nature. 2008 Oct 16;455(7215):879-80. doi: 10.1038/455879a. Nature. 2008. PMID: 18923500 No abstract available.
Similar articles
-
Conformational flexibility and peptide interaction of the translocation ATPase SecA.J Mol Biol. 2009 Dec 11;394(4):606-12. doi: 10.1016/j.jmb.2009.10.024. Epub 2009 Oct 20. J Mol Biol. 2009. PMID: 19850053 Free PMC article.
-
Conformational Changes of the Clamp of the Protein Translocation ATPase SecA.J Mol Biol. 2015 Jul 17;427(14):2348-59. doi: 10.1016/j.jmb.2015.05.003. Epub 2015 May 14. J Mol Biol. 2015. PMID: 25982945 Free PMC article.
-
Crystal structure of a substrate-engaged SecY protein-translocation channel.Nature. 2016 Mar 17;531(7594):395-399. doi: 10.1038/nature17163. Epub 2016 Mar 7. Nature. 2016. PMID: 26950603 Free PMC article.
-
[Molecular mechanisms of SecA-mediated protein translocation viewed from structural studies].Tanpakushitsu Kakusan Koso. 2009 May;54(6):685-95. Tanpakushitsu Kakusan Koso. 2009. PMID: 19462754 Review. Japanese. No abstract available.
-
Oligomeric states of the SecA and SecYEG core components of the bacterial Sec translocon.Biochim Biophys Acta. 2007 Jan;1768(1):5-12. doi: 10.1016/j.bbamem.2006.08.013. Epub 2006 Aug 30. Biochim Biophys Acta. 2007. PMID: 17011510 Free PMC article. Review.
Cited by
-
Comparison of Single and Multiple Turnovers of SecYEG in Escherichia coli.J Bacteriol. 2020 Nov 19;202(24):e00462-20. doi: 10.1128/JB.00462-20. Print 2020 Nov 19. J Bacteriol. 2020. PMID: 32989086 Free PMC article.
-
The Structure of Clostridioides difficile SecA2 ATPase Exposes Regions Responsible for Differential Target Recognition of the SecA1 and SecA2-Dependent Systems.Int J Mol Sci. 2020 Aug 26;21(17):6153. doi: 10.3390/ijms21176153. Int J Mol Sci. 2020. PMID: 32858965 Free PMC article.
-
Cotranslational folding of alkaline phosphatase in the periplasm of Escherichia coli.Protein Sci. 2020 Oct;29(10):2028-2037. doi: 10.1002/pro.3927. Epub 2020 Aug 24. Protein Sci. 2020. PMID: 32790204 Free PMC article.
-
Identification of small-molecule inhibitors against SecA by structure-based virtual ligand screening.J Antibiot (Tokyo). 2015 Nov;68(11):666-73. doi: 10.1038/ja.2015.53. Epub 2015 May 20. J Antibiot (Tokyo). 2015. PMID: 25990955
-
Protein transport across and into cell membranes in bacteria and archaea.Cell Mol Life Sci. 2010 Jan;67(2):179-99. doi: 10.1007/s00018-009-0160-x. Epub 2009 Oct 10. Cell Mol Life Sci. 2010. PMID: 19823765 Free PMC article. Review.
References
-
- Rapoport TA Protein translocation across the eukaryotic endoplasmic reticulum and bacterial plasma membranes. Nature 450, 663–669 (2007). - PubMed
-
- Qi HY & Bernstein HD SecA is required for the insertion of inner membrane proteins targeted by the Escherichia coli signal recognition particle. J Biol Chem 274, 8993–8997 (1999). - PubMed
-
- van den Berg B et al. X-ray structure of a protein-conducting channel. Nature 427, 36–44 (2004). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
