

A key role for autophagy and the autophagy gene Atg16l1 in mouse and human intestinal Paneth cells
- PMID: 18849966
- PMCID: PMC2695978
- DOI: 10.1038/nature07416
A key role for autophagy and the autophagy gene Atg16l1 in mouse and human intestinal Paneth cells
Abstract
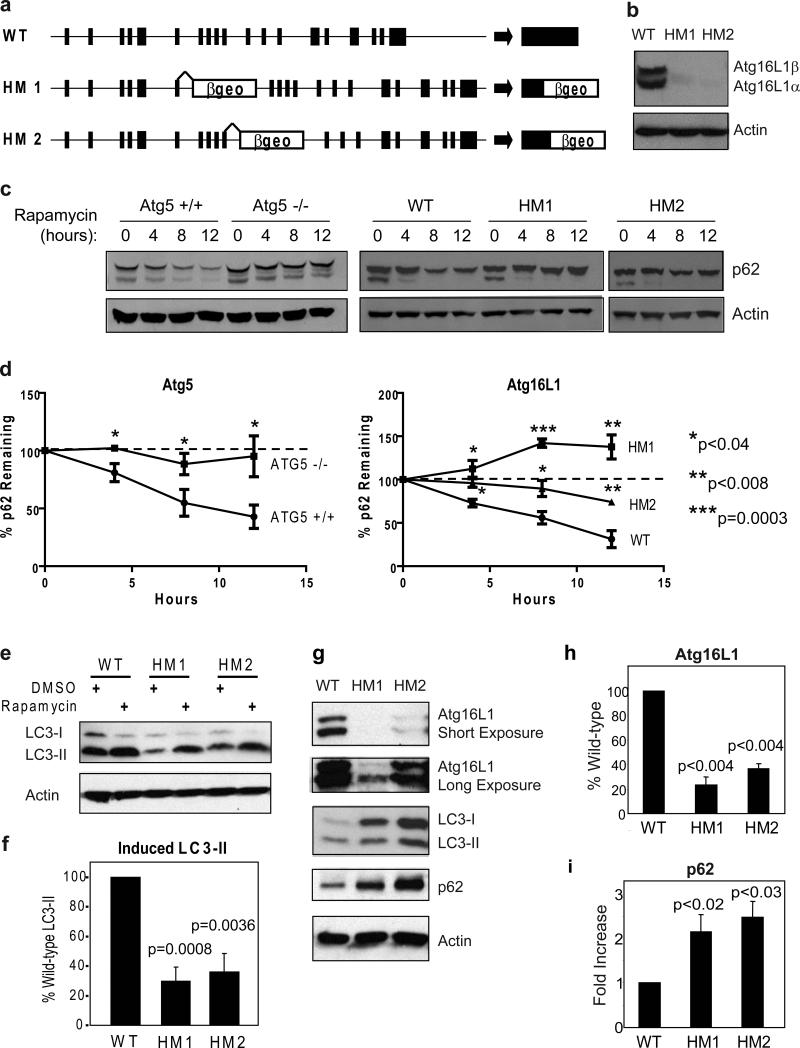
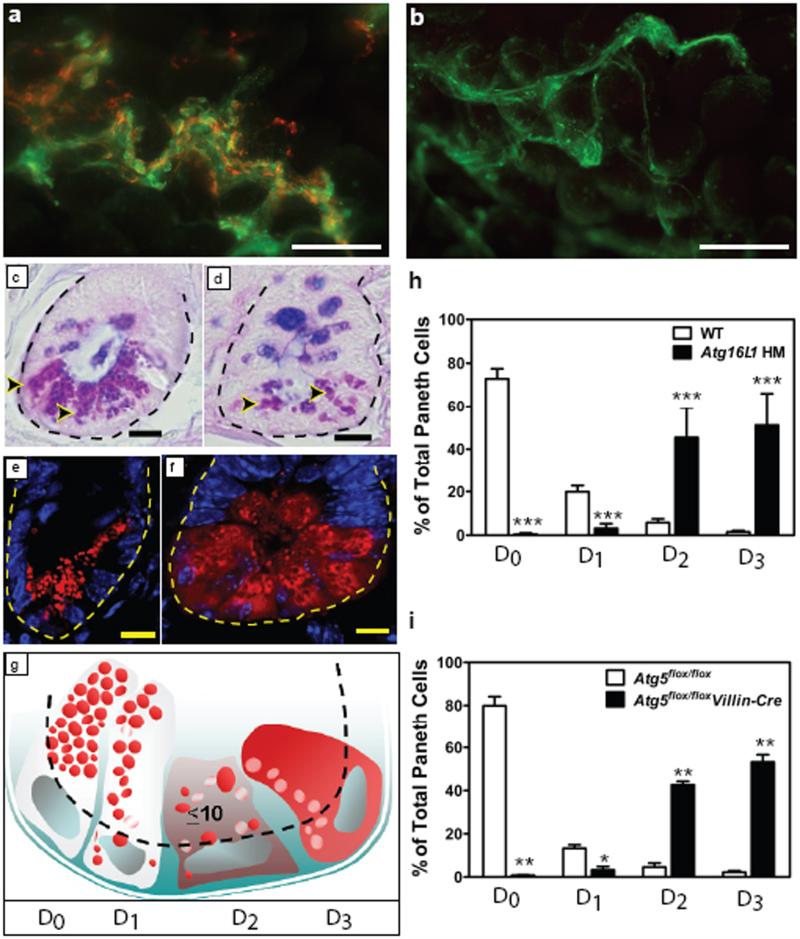
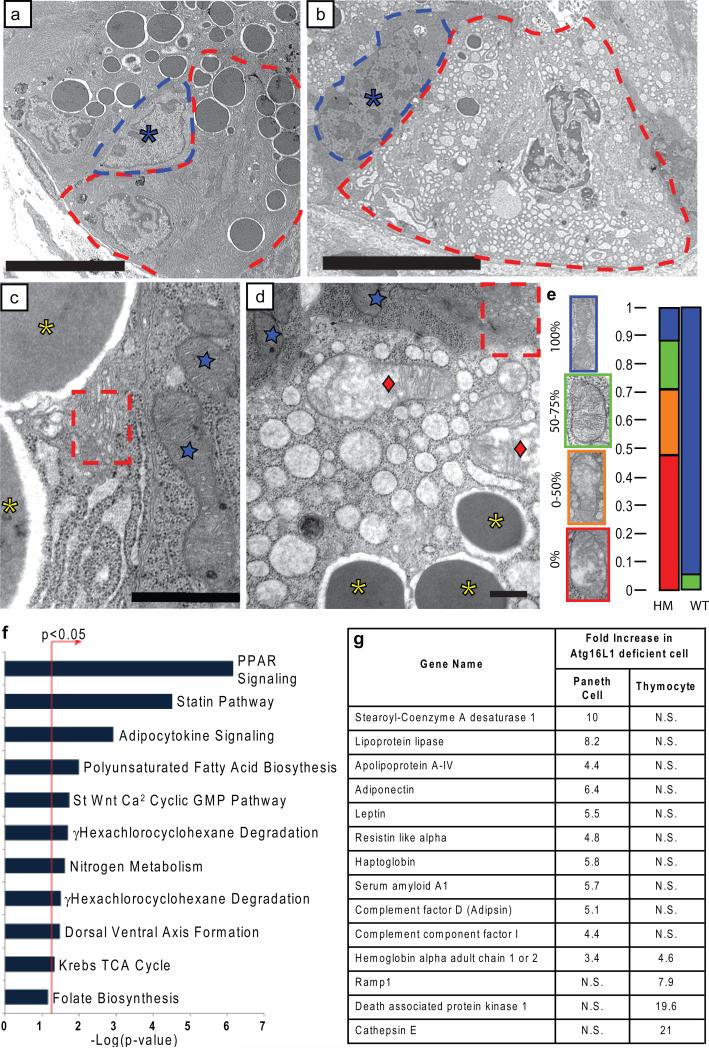
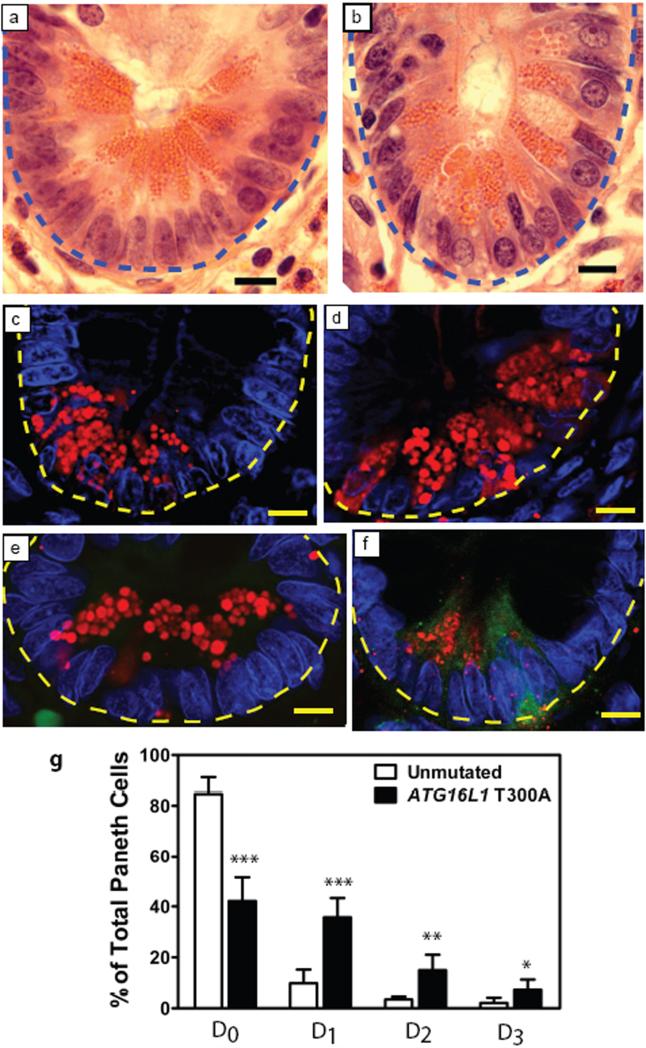
Susceptibility to Crohn's disease, a complex inflammatory disease involving the small intestine, is controlled by over 30 loci. One Crohn's disease risk allele is in ATG16L1, a gene homologous to the essential yeast autophagy gene ATG16 (ref. 2). It is not known how ATG16L1 or autophagy contributes to intestinal biology or Crohn's disease pathogenesis. To address these questions, we generated and characterized mice that are hypomorphic for ATG16L1 protein expression, and validated conclusions on the basis of studies in these mice by analysing intestinal tissues that we collected from Crohn's disease patients carrying the Crohn's disease risk allele of ATG16L1. Here we show that ATG16L1 is a bona fide autophagy protein. Within the ileal epithelium, both ATG16L1 and a second essential autophagy protein ATG5 are selectively important for the biology of the Paneth cell, a specialized epithelial cell that functions in part by secretion of granule contents containing antimicrobial peptides and other proteins that alter the intestinal environment. ATG16L1- and ATG5-deficient Paneth cells exhibited notable abnormalities in the granule exocytosis pathway. In addition, transcriptional analysis revealed an unexpected gain of function specific to ATG16L1-deficient Paneth cells including increased expression of genes involved in peroxisome proliferator-activated receptor (PPAR) signalling and lipid metabolism, of acute phase reactants and of two adipocytokines, leptin and adiponectin, known to directly influence intestinal injury responses. Importantly, Crohn's disease patients homozygous for the ATG16L1 Crohn's disease risk allele displayed Paneth cell granule abnormalities similar to those observed in autophagy-protein-deficient mice and expressed increased levels of leptin protein. Thus, ATG16L1, and probably the process of autophagy, have a role within the intestinal epithelium of mice and Crohn's disease patients by selective effects on the cell biology and specialized regulatory properties of Paneth cells.
Figures
Comment in
-
Defects in autophagy induce alterations in the secretory pathway and proinflammatory signaling of paneth cells.Gastroenterology. 2009 Oct;137(4):1527-9. doi: 10.1053/j.gastro.2009.08.021. Epub 2009 Aug 28. Gastroenterology. 2009. PMID: 19717131 No abstract available.
Similar articles
-
A common role for Atg16L1, Atg5 and Atg7 in small intestinal Paneth cells and Crohn disease.Autophagy. 2009 Feb;5(2):250-2. doi: 10.4161/auto.5.2.7560. Epub 2009 Feb 8. Autophagy. 2009. PMID: 19139628 Free PMC article.
-
Genomic ATG16L1 risk allele-restricted Paneth cell ER stress in quiescent Crohn's disease.Gut. 2014 Jul;63(7):1081-91. doi: 10.1136/gutjnl-2012-303527. Epub 2013 Aug 20. Gut. 2014. PMID: 23964099
-
Paneth cells as a site of origin for intestinal inflammation.Nature. 2013 Nov 14;503(7475):272-6. doi: 10.1038/nature12599. Epub 2013 Oct 2. Nature. 2013. PMID: 24089213 Free PMC article.
-
Classification of genetic profiles of Crohn's disease: a focus on the ATG16L1 gene.Expert Rev Mol Diagn. 2008 Mar;8(2):199-207. doi: 10.1586/14737159.8.2.199. Expert Rev Mol Diagn. 2008. PMID: 18366306 Review.
-
[Barrier- and autophagic functions of the intestinal epithelia: role of disturbances in the pathogenesis of Crohn's disease].Orv Hetil. 2010 Oct 3;151(40):1645-55. doi: 10.1556/OH.2010.28942. Orv Hetil. 2010. PMID: 20860961 Review. Hungarian.
Cited by
-
Toll-like receptor 7 protects against intestinal inflammation and restricts the development of colonic tissue-resident memory CD8+ T cells.Front Immunol. 2024 Oct 11;15:1465175. doi: 10.3389/fimmu.2024.1465175. eCollection 2024. Front Immunol. 2024. PMID: 39464882 Free PMC article.
-
Persistent Activation of the P2X7 Receptor Underlies Chronic Inflammation and Carcinogenic Changes in the Intestine.Int J Mol Sci. 2024 Oct 10;25(20):10874. doi: 10.3390/ijms252010874. Int J Mol Sci. 2024. PMID: 39456655 Free PMC article. Review.
-
Host-pathobiont interactions in Crohn's disease.Nat Rev Gastroenterol Hepatol. 2024 Oct 24. doi: 10.1038/s41575-024-00997-y. Online ahead of print. Nat Rev Gastroenterol Hepatol. 2024. PMID: 39448837 Review.
-
Zebrafish reveal new roles for Fam83f in hatching and the DNA damage-mediated autophagic response.Open Biol. 2024 Oct;14(10):240194. doi: 10.1098/rsob.240194. Epub 2024 Oct 23. Open Biol. 2024. PMID: 39437839 Free PMC article.
-
Complement C3 deposition restricts the proliferation of internalized Staphylococcus aureus by promoting autophagy.Front Cell Infect Microbiol. 2024 Sep 6;14:1400068. doi: 10.3389/fcimb.2024.1400068. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 39310788 Free PMC article.
References
-
- Mizushima N, et al. Mouse Apg16L, a novel WD-repeat protein, targets to the autophagic isolation membrane with the Apg12-Apg5 conjugate. J. Cell Sci. 2003;116:1679–1688. - PubMed
-
- Ouellette AJ. Paneth cell alpha-defensin synthesis and function. Curr. Top. Microbiol. Immunol. 2006;306:1–25. - PubMed
-
- Barbier M, et al. Overexpression of leptin mRNA in mesenteric adipose tissue in inflammatory bowel diseases. Gastroenterol. Clin. Biol. 2003;27:987–991. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
- AI062773/AI/NIAID NIH HHS/United States
- P30 DK040561/DK/NIDDK NIH HHS/United States
- T32 AR007279-30/AR/NIAMS NIH HHS/United States
- P30 DK043351/DK/NIDDK NIH HHS/United States
- T32 AR007279/AR/NIAMS NIH HHS/United States
- DK43351/DK/NIDDK NIH HHS/United States
- U54 AI057160-010005/AI/NIAID NIH HHS/United States
- P30 DK052574-09/DK/NIDDK NIH HHS/United States
- R01 AI062832-04/AI/NIAID NIH HHS/United States
- U54 AI057160-05S10018/AI/NIAID NIH HHS/United States
- R01 AI062832/AI/NIAID NIH HHS/United States
- P30 DK043351-18/DK/NIDDK NIH HHS/United States
- R01 AI062773-01A1/AI/NIAID NIH HHS/United States
- R01 AI062773/AI/NIAID NIH HHS/United States
- P30 DK52574/DK/NIDDK NIH HHS/United States
- U54 AI057160/AI/NIAID NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
- T32 AR07279/AR/NIAMS NIH HHS/United States
- P30 DK040561-13/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
