

Insights into interferon regulatory factor activation from the crystal structure of dimeric IRF5
- PMID: 18836453
- PMCID: PMC2757928
- DOI: 10.1038/nsmb.1496
Insights into interferon regulatory factor activation from the crystal structure of dimeric IRF5
Abstract
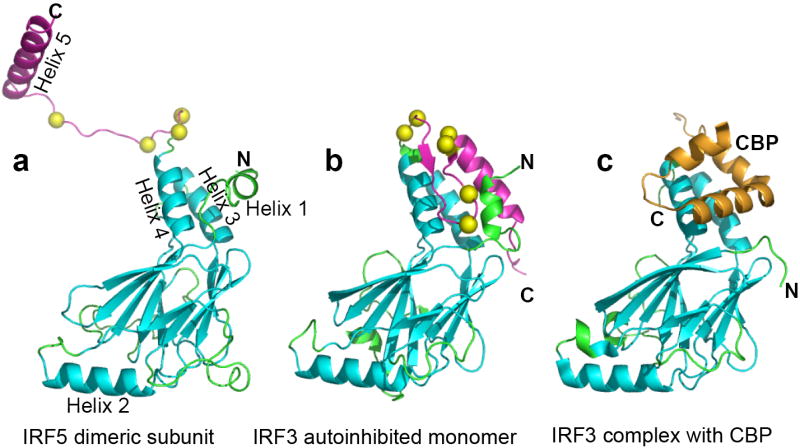
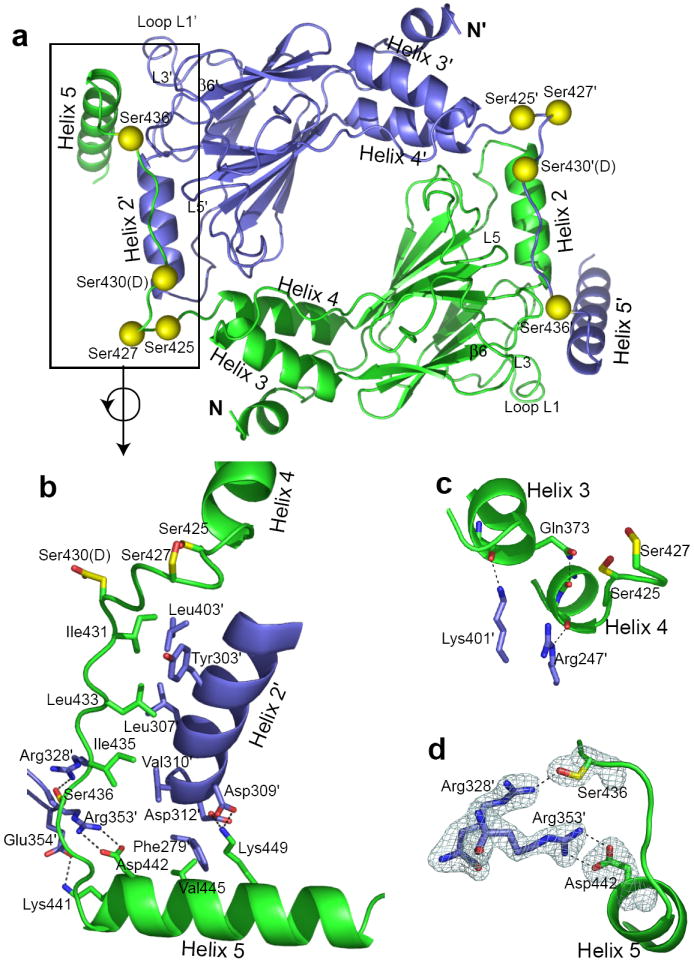
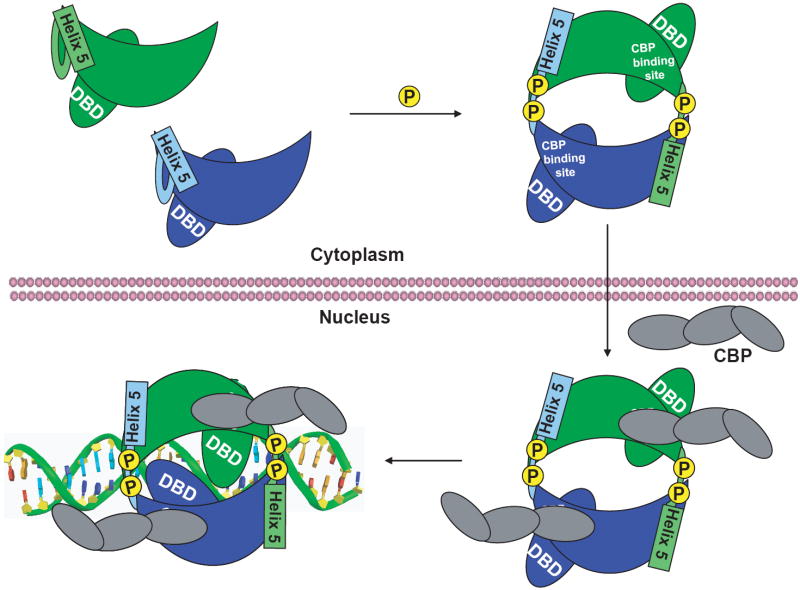
Interferon regulatory factors (IRFs) are essential in the innate immune response and other physiological processes. Activation of these proteins in the cytoplasm is triggered by phosphorylation of serine and threonine residues in a C-terminal autoinhibitory region, which stimulates dimerization, transport into the nucleus, assembly with the coactivator CBP/p300 and initiation of transcription. The crystal structure of the transactivation domain of pseudophosphorylated human IRF5 strikingly reveals a dimer in which the bulk of intersubunit interactions involve a highly extended C-terminal region. The corresponding region has previously been shown to block CBP/p300 binding to unphosphorylated IRF3. Mutation of key interface residues supports the observed dimer as the physiologically activated state of IRF5 and IRF3. Thus, phosphorylation is likely to activate IRF5 and other family members by triggering conformational rearrangements that switch the C-terminal segment from an autoinihibitory to a dimerization role.
Figures

Similar articles
-
Advances and challenges in targeting IRF5, a key regulator of inflammation.FEBS J. 2019 May;286(9):1624-1637. doi: 10.1111/febs.14654. Epub 2018 Sep 21. FEBS J. 2019. PMID: 30199605 Free PMC article. Review.
-
Structural insights into interferon regulatory factor activation.Cell Signal. 2010 Jun;22(6):883-7. doi: 10.1016/j.cellsig.2009.12.005. Epub 2009 Dec 30. Cell Signal. 2010. PMID: 20043992 Free PMC article. Review.
-
Rotavirus NSP1 mediates degradation of interferon regulatory factors through targeting of the dimerization domain.J Virol. 2013 Sep;87(17):9813-21. doi: 10.1128/JVI.01146-13. Epub 2013 Jul 3. J Virol. 2013. PMID: 23824805 Free PMC article.
-
Molecular characterization and expression analysis of eleven interferon regulatory factors in half-smooth tongue sole, Cynoglossus semilaevis.Fish Shellfish Immunol. 2015 May;44(1):272-82. doi: 10.1016/j.fsi.2015.02.033. Epub 2015 Feb 28. Fish Shellfish Immunol. 2015. PMID: 25731919
-
Activation of interferon regulatory factor 5 by site specific phosphorylation.PLoS One. 2012;7(3):e33098. doi: 10.1371/journal.pone.0033098. Epub 2012 Mar 8. PLoS One. 2012. PMID: 22412986 Free PMC article.
Cited by
-
Pestivirus Npro Directly Interacts with Interferon Regulatory Factor 3 Monomer and Dimer.J Virol. 2016 Aug 12;90(17):7740-7. doi: 10.1128/JVI.00318-16. Print 2016 Sep 1. J Virol. 2016. PMID: 27334592 Free PMC article.
-
A conserved region within interferon regulatory factor 5 controls breast cancer cell migration through a cytoplasmic and transcription-independent mechanism.Mol Cancer. 2015 Feb 4;14(1):32. doi: 10.1186/s12943-015-0305-5. Mol Cancer. 2015. PMID: 25649192 Free PMC article.
-
An IRF5 Decoy Peptide Reduces Myocardial Inflammation and Fibrosis and Improves Endothelial Cell Function in Tight-Skin Mice.PLoS One. 2016 Apr 6;11(4):e0151999. doi: 10.1371/journal.pone.0151999. eCollection 2016. PLoS One. 2016. PMID: 27050551 Free PMC article.
-
Interferon regulatory factor 5 in the pathogenesis of systemic lupus erythematosus.Clin Dev Immunol. 2012;2012:780436. doi: 10.1155/2012/780436. Epub 2012 Nov 1. Clin Dev Immunol. 2012. PMID: 23251221 Free PMC article. Review.
-
Advances and challenges in targeting IRF5, a key regulator of inflammation.FEBS J. 2019 May;286(9):1624-1637. doi: 10.1111/febs.14654. Epub 2018 Sep 21. FEBS J. 2019. PMID: 30199605 Free PMC article. Review.
References
-
- Honda K, Taniguchi T. IRFs: master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat Rev Immunol. 2006;6:644–58. - PubMed
-
- Tamura T, Yanai H, Savitsky D, Taniguchi T. The IRF Family Transcription Factors in Immunity and Oncogenesis. Annu Rev Immunol. 2008;26:535–84. - PubMed
-
- Schoenemeyer A, et al. The interferon regulatory factor, IRF5, is a central mediator of toll-like receptor 7 signaling. J Biol Chem. 2005;280:17005–12. - PubMed
-
- Yasuda K, et al. Murine dendritic cell type I IFN production induced by human IgG-RNA immune complexes is IFN regulatory factor (IRF)5 and IRF7 dependent and is required for IL-6 production. J Immunol. 2007;178:6876–85. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
