

JAK2/STAT2/STAT3 are required for myogenic differentiation
- PMID: 18835816
- PMCID: PMC2662224
- DOI: 10.1074/jbc.M803012200
JAK2/STAT2/STAT3 are required for myogenic differentiation
Abstract
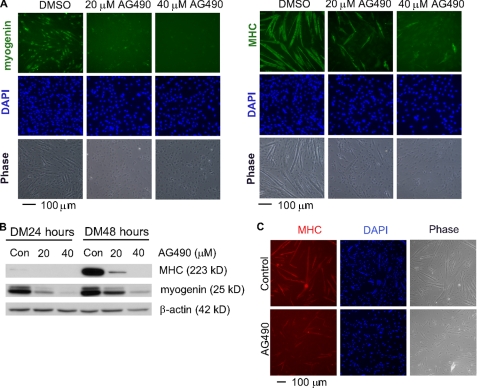
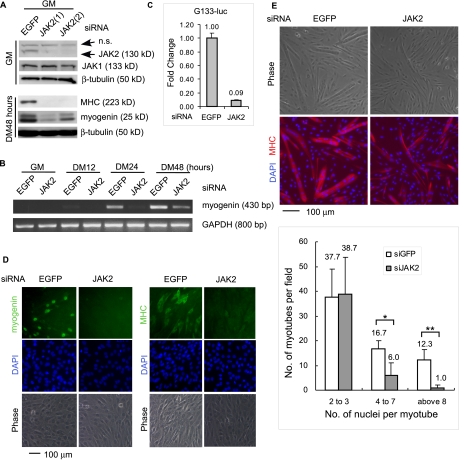
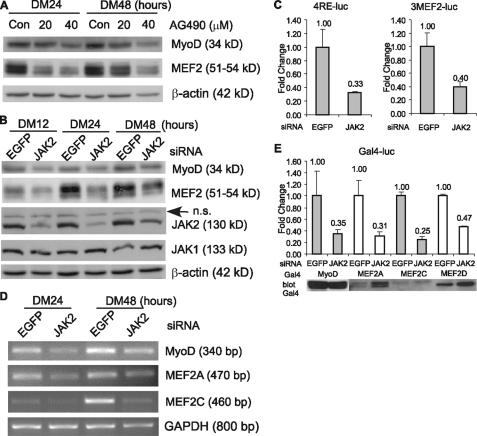
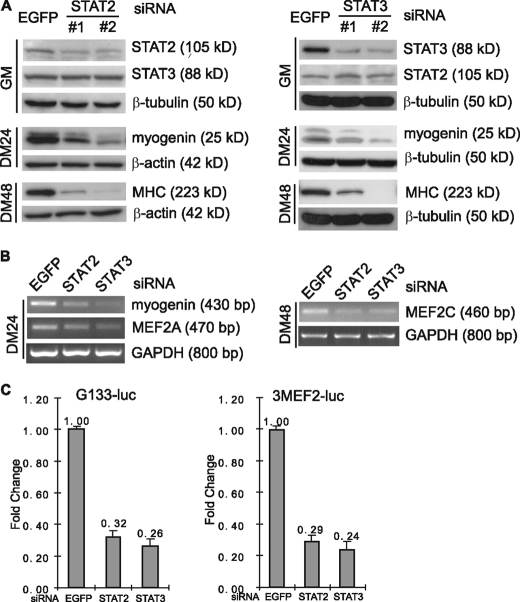
Skeletal muscle satellite cell-derived myoblasts are mainly responsible for postnatal muscle growth and injury-induced regeneration. However, the cellular signaling pathways that control proliferation and differentiation of myoblasts remain poorly defined. Recently, we found that JAK1/STAT1/STAT3 not only participate in myoblast proliferation but also actively prevent them from premature differentiation. Unexpectedly, we found that a related pathway consisting of JAK2, STAT2, and STAT3 is required for early myogenic differentiation. Interference of this pathway by either a small molecule inhibitor or small interfering RNA inhibits myogenic differentiation. Consistently, all three molecules are activated upon differentiation. The pro-differentiation effect of JAK2/STAT2/STAT3 is partially mediated by MyoD and MEF2. Interestingly, the expression of the IGF2 gene and the HGF gene is also regulated by JAK2/STAT2/STAT3, suggesting that this pathway could also promote differentiation by regulating signaling molecules known to be involved in myogenic differentiation. In summary, our current study reveals a novel role for the JAK2/STAT2/STAT3 pathway in myogenic differentiation.
Figures
Similar articles
-
JAK1-STAT1-STAT3, a key pathway promoting proliferation and preventing premature differentiation of myoblasts.J Cell Biol. 2007 Oct 8;179(1):129-38. doi: 10.1083/jcb.200703184. Epub 2007 Oct 1. J Cell Biol. 2007. PMID: 17908914 Free PMC article.
-
Interleukin-6 Induces Myogenic Differentiation via JAK2-STAT3 Signaling in Mouse C2C12 Myoblast Cell Line and Primary Human Myoblasts.Int J Mol Sci. 2019 Oct 24;20(21):5273. doi: 10.3390/ijms20215273. Int J Mol Sci. 2019. PMID: 31652937 Free PMC article.
-
Role of JAK3 in myogenic differentiation.Cell Signal. 2012 Mar;24(3):742-9. doi: 10.1016/j.cellsig.2011.11.009. Epub 2011 Nov 18. Cell Signal. 2012. PMID: 22120524
-
JAK-STAT pathway and myogenic differentiation.JAKSTAT. 2013 Apr 1;2(2):e23282. doi: 10.4161/jkst.23282. JAKSTAT. 2013. PMID: 24058805 Free PMC article. Review.
-
STAT3 in Skeletal Muscle Function and Disorders.Int J Mol Sci. 2018 Aug 2;19(8):2265. doi: 10.3390/ijms19082265. Int J Mol Sci. 2018. PMID: 30072615 Free PMC article. Review.
Cited by
-
Transcriptome Response of Differentiating Muscle Satellite Cells to Thermal Challenge in Commercial Turkey.Genes (Basel). 2022 Oct 14;13(10):1857. doi: 10.3390/genes13101857. Genes (Basel). 2022. PMID: 36292741 Free PMC article.
-
STAT3 promotes IFNγ/TNFα-induced muscle wasting in an NF-κB-dependent and IL-6-independent manner.EMBO Mol Med. 2017 May;9(5):622-637. doi: 10.15252/emmm.201607052. EMBO Mol Med. 2017. PMID: 28264935 Free PMC article.
-
Evidence of a broad histamine footprint on the human exercise transcriptome.J Physiol. 2016 Sep 1;594(17):5009-23. doi: 10.1113/JP272177. Epub 2016 May 29. J Physiol. 2016. PMID: 27061420 Free PMC article. Clinical Trial.
-
The E3 ubiquitin ligase Nedd4L preserves skeletal muscle stem cell quiescence by inhibiting their activation.iScience. 2024 Jun 11;27(7):110241. doi: 10.1016/j.isci.2024.110241. eCollection 2024 Jul 19. iScience. 2024. PMID: 39015146 Free PMC article.
-
Chromatin signaling in muscle stem cells: interpreting the regenerative microenvironment.Front Aging Neurosci. 2015 Apr 7;7:36. doi: 10.3389/fnagi.2015.00036. eCollection 2015. Front Aging Neurosci. 2015. PMID: 25904863 Free PMC article. Review.
References
-
- Molkentin, J. D., and Olson, E. N. (1996) Curr. Opin. Genet. Dev. 6 445-453 - PubMed
-
- Sabourin, L. A., and Rudnicki, M. A. (2000) Clin. Genet. 57 16-25 - PubMed
-
- Tapscott, S. J. (2005) Development 132 2685-2695 - PubMed
-
- Black, B. L., and Olson, E. N. (1998) Annu. Rev. Cell Dev. Biol. 14 167-196 - PubMed
-
- McKinsey, T. A., Zhang, C. L., and Olson, E. N. (2002) Trends Biochem. Sci. 27 40-47 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
