

Review
doi: 10.1016/j.molcel.2008.07.002.
Lysine acetylation: codified crosstalk with other posttranslational modifications
Affiliations
- PMID: 18722172
- PMCID: PMC2551738
- DOI: 10.1016/j.molcel.2008.07.002
Item in Clipboard
Review
Lysine acetylation: codified crosstalk with other posttranslational modifications
Mol Cell.
.
Abstract
Lysine acetylation has emerged as a major posttranslational modification for histones. Crossregulation between this and other modifications is crucial in modulating chromatin-based transcriptional control and shaping inheritable epigenetic programs. In addition to histones, many other nuclear proteins and various cytoplasmic regulators are subject to lysine acetylation. This review focuses on recent findings pertinent to acetylation of nonhistone proteins and emphasizes how this modification might crosstalk with phosphorylation, methylation, ubiquitination, sumoylation, and others to form code-like multisite modification programs for dynamic control of cellular signaling under diverse conditions.
Figures
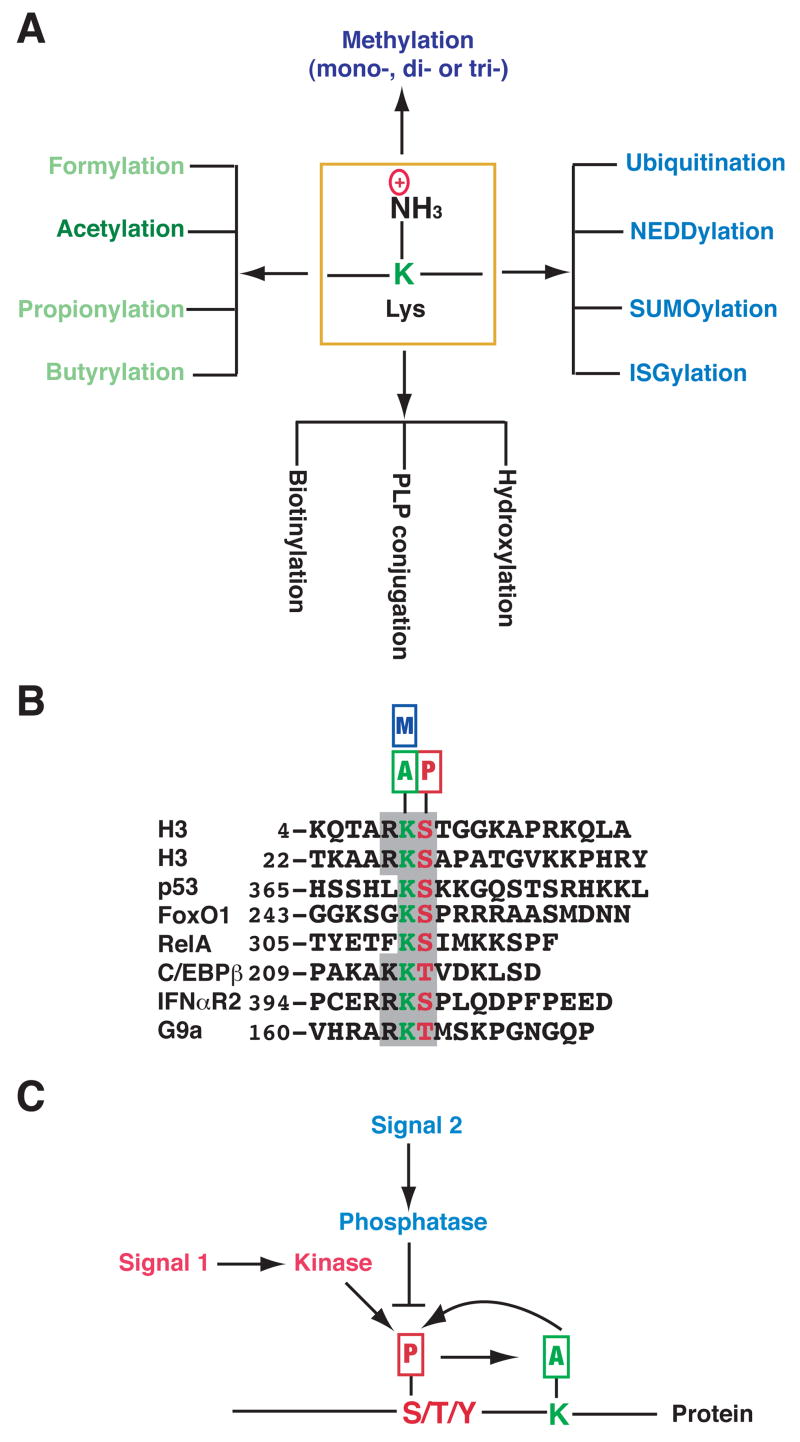
(A) The side chain of a Lys (K) residue can be modified by acetylation and other covalent modifications. Except for hydroxylation, all listed modifications target the ε-amino group. Formylation, propionylation and butyrylation are three recently discovered modifications that are structurally similar to acetylation; however, it is not known if formylation occurs enzymatically. Aside from formylation, hydroxylation and PLP (pyridoxal 5′-phosphate, a vitamin B6 derivative) conjugation, all listed modifications are reversible. Acetylation of a Lys residue precludes further modifications by others, and vice versa. (B) Alignment of sequences surrounding KS dipeptides from histone H3, p53 and several other proteins. The KS dipeptides are highlighted in color. Small rectangles with the letters A, P and M denote acetylation, phosphorylation and methylation, respectively. IFNαR2 (interferon α receptor 2) acetylation at Lys 399 and phosphorylation at Ser 400 might crosstalk with one another (Tang et al., 2007). It remains to be established whether similar interplay occurs in p53, FoxO1, RelA (NF-κB subunit) (Perkins, 2006), and C/EBPβ (CCAAT/enhancer binding protein β) (Hasselgren, 2007). Methylation of the highlighted Lys residues has been documented only for histone H3, p53 (Huang et al., 2006b) and G9a (Sampath et al., 2007). It is unclear whether the Ser in p53 is phosphorylated or the Lys in G9a is acetylated. (C) Cartoon illustrating crosstalk between phosphorylation and acetylation. Different signals act on Ser (S), Thr (T) or Tyr (Y) phosphorylation, which in turn affects acetylation of a neighboring Lys. Acetylation might also regulate phosphorylation. The Lys can be adjacent to or far away from the phosphorylation site, which can be either N-terminal or C-terminal from the acetylation site.
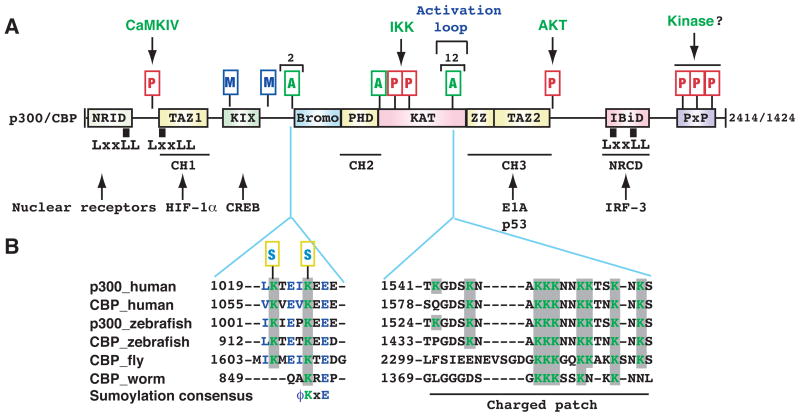
(A) Domain organization of human p300 and CBP. As mammalian p300 and CBP are functionally interchangeable in many in vitro assays, the term p300/CBP is used to refer to one or the other. Acetylation, phosphorylation, methylation and sumoylation sites are denoted with small rectangles containing the letters A, P, M and S, respectively. Within the activation loop of p300 and perhaps also CBP, there are 12 acetylation sites. The responsible kinases are indicated for the phosphorylation sites. Representative transcription factors that use p300 and CBP as coactivators are listed along with arrows pointing to the respective binding domains. Domain abbreviation: Bromo, bromodomain; CH1, Cys- & His-rich domain 1; IBiD, IRF-3 binding domain; KIX, kinase-inducing domain (KID) binding region; NRCD, nuclear receptor coactivator binding domain (synonymous to IBiD); NRID, nuclear receptor interacting domain; PxP, Pro-rich domain (no known function); TAZ1, transcriptional adaptor zinc finger; ZZ, zinc finger near the dystrophin WW domain. (B) Regional sequence comparison of p300 and CBP from human and other species. The alignment was generated and shaded using ClustalW of MacVector 7.2 (Accelrys), using GenBank accession numbers Q09472, 119943104, 125830995, 125850731, 24640865, and 17552710. According to the new nomenclature system for histone-modifying enzymes (Allis et al., 2007), mammalian CBP and p300 are referred to as KAT3A and KAT3B, respectively. In the sumoylation consensus, φ is preferably a bulky residue (e.g., Leu, Ile or Val). Note that Lys 1057 of mouse CBP (1056 in humans) has been shown to be sumoylated (Kuo et al., 2005).
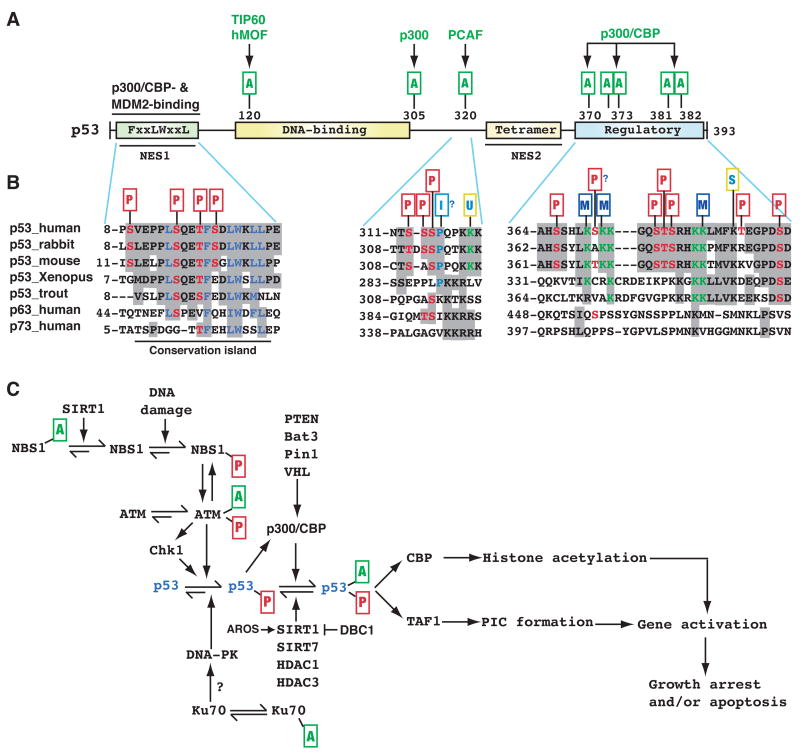
(A) Domain organization of human p53. Residues 8–28 harbor overlapping binding sites for p300/CBP and the ubiquitin ligase MDM2, whereas within residues 369–393 there is a cluster of five Lys residues acetylated by p300/CBP. These residues are also targets of ubiquitination and neddylation (Brooks and Gu, 2003). Other acetylation sites are shown along with the responsible KATs. Unlike Lys372, methylation of Lys 370 or 382 inhibits p53 activity. NES, nuclear export signal; tetramer, tetramerization domain; regulatory, regulatory region. (B) Sequences encompassing residues 8–28, 311–321 and 369–393 of human p53 are compared with the corresponding regions of orthologs and paralogs. p53 contains numerous phosphorylation sites (Toledo and Wahl, 2006), but only those located within these three regions are shown. PTMs are marked as in Fig. 2, and the small rectangle with letter I denotes proline isomerization. Sequence analysis was performed as in Fig. 2B, using GenBank accession numbers ABM86630, 126722898, 148747262, NP_0010811568, P25035, 31543818, and 4885645. Residues 10–27 are highly conserved from zebrafish to humans and are also more conserved than the flanking sequences, so this region forms a small ‘conservation island’ (Yang and Grégoire, 2006). In comparison, residues 311–321 and 369–393 are less conserved. (C) In DNA damage responses, Lys acetylation targets not only p53 itself but also various components of the signaling pathways that control p53 activation. Exactly how acetylation of upstream components affect p53 regulation remains to be established. AROS, active regulator of SIRT1; Bat3, HLA B-associated transcript 3 (also known as Scythe); DBC1 (deleted in breast cancer 1); PIC, transcriptional pre-initiation complex; Pin1, protein interacting with NIMA (never in mitosis A)-1; TAF1, TBP-associated factor 1; VHL, von Hippel-Lindau gene product. See text for more details.
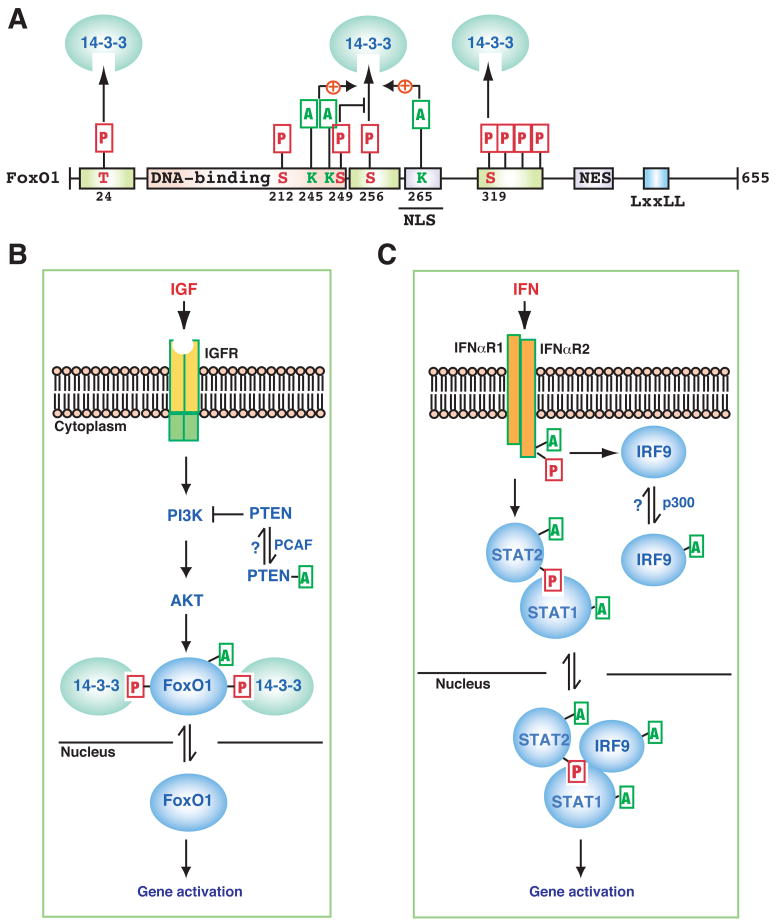
(A) Domain organization of human FoxO1. In addition to a highly conserved DNA-binding domain in the middle, FoxO1 possesses six conserved motifs, three of which are phosphorylated for 14-3-3 binding and nuclear export. The critical Thr (T) and Ser (S) residues are indicated along with their positions. In addition, a nuclear localization signal (NLS), a nuclear export signal (NES), and an LxxLL motif are conserved. (B–C) Schematics showing how Lys acetylation modulates pathways that transduce signals from insulin-like growth factor (IGF, B) and interferon (IFN, C) to the nucleus. IGFR, IGF receptor; IFNαR1, IFNα receptor 1. See text for details.
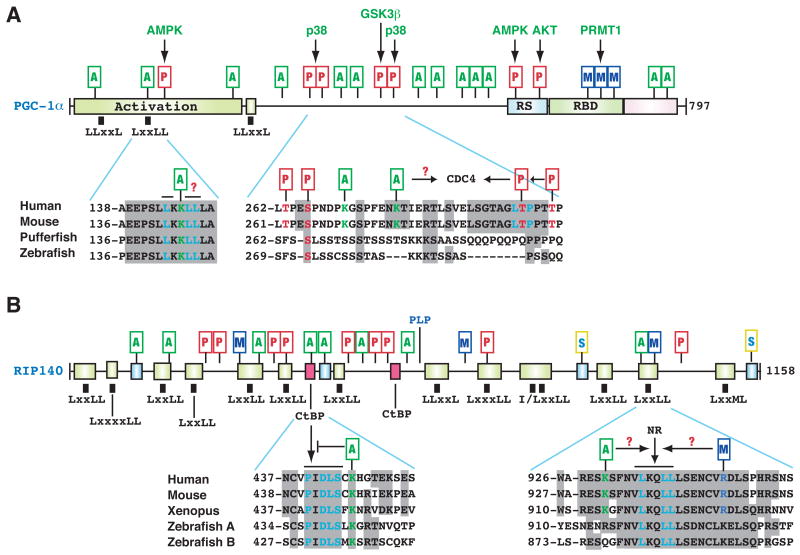
(A) Domain organization of human PGC-1α, with conserved domains shown in colored boxes. One LxxLL and two LLxxL motifs are depicted by small black boxes. These motifs are evolutionarily conserved. Within the LxxLL motif, the acetylatable Lys is highlighted in green. PTMs are maked as in Fig. 2. Activation, transcriptional activation domain; RS, Arg- and Ser-rich domain; RBD, RNA-binding domain. (B) Domain organization of human RIP140. Conserved domains are shown in small colored boxes. Small black boxes indicate the rough positions of Leu-rich motifs. RIP140 contains two conserved CtBP-binding motifs and double sumoylation sites. NR, nuclear receptor. Sequence analyses were performed as in Fig. 2B, using GenBank accession numbers 7019499, AAH66868, CAG02304, and AAY15212 (for PGC-1α); CAA59108, NP_032761, NP_001083708, 125838185, and 125828075 (for RIP140). See text for details.
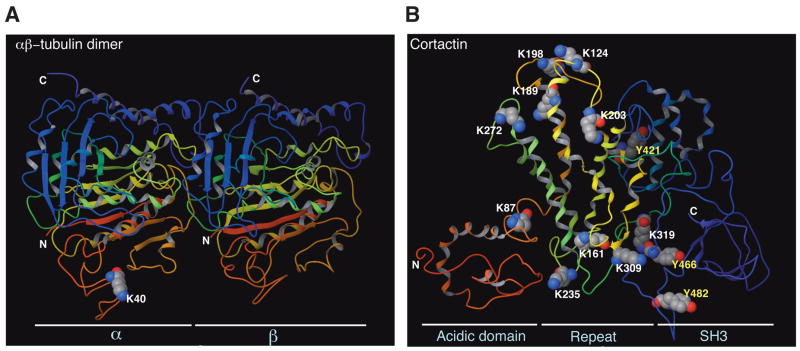
(A) Ribbon representation of an α/β-tubulin heterodimer. Electron diffraction data was obtained from Protein Data Bank (PDB ID 1tub) for preparation of the 3D structure using the Maestro program (version 8). N and C mark the N- and C-terminal ends, respectively. Whereas Lys 40 of α-tubulin faces the lumenal side, the C-terminal ends of α- and β-tubulin are outside from microtubules. (B) Ribbon representation of the cortactin structure. Electron diffraction data was obtained from Protein Data Bank (PDB ID 2F9X). The 3D structure was generated along with acetylation sites and three phospho-Tyr residues colored as in (A). Lys 161, 235, 309 and 319 form one charged patch, whereas Lys 124, 189, 198, 203, and 272 form another. Albeit over 200 residues apart in the primary sequence, Lys 203 and Tyr 421 are close to one another at the 3D level. Similarly, Lys 309 and 319 are adjacent to Tyr 466 in the 3D structure. Physical proximity at the 3D level suggests potential crosstalk between Lys acetylation and Tyr phosphorylation, possibly linking cellular signaling to regulation of reversible acetylation.
Similar articles
-
Crosstalk between CARM1 methylation and CBP acetylation on histone H3.Curr Biol. 2002 Dec 23;12(24):2090-7. doi: 10.1016/s0960-9822(02)01387-8. Curr Biol. 2002. PMID: 12498683
-
Lysine acetylation: enzymes, bromodomains and links to different diseases.Essays Biochem. 2012;52:1-12. doi: 10.1042/bse0520001. Essays Biochem. 2012. PMID: 22708559 Review.
-
Lysine methylation: beyond histones.Acta Biochim Biophys Sin (Shanghai). 2012 Jan;44(1):14-27. doi: 10.1093/abbs/gmr100. Acta Biochim Biophys Sin (Shanghai). 2012. PMID: 22194010 Review.
-
Aberrant lysine acetylation in tumorigenesis: Implications in the development of therapeutics.Pharmacol Ther. 2016 Jun;162:98-119. doi: 10.1016/j.pharmthera.2016.01.011. Epub 2016 Jan 22. Pharmacol Ther. 2016. PMID: 26808162 Review.
-
Beyond histones - the expanding roles of protein lysine methylation.FEBS J. 2017 Sep;284(17):2732-2744. doi: 10.1111/febs.14056. Epub 2017 May 2. FEBS J. 2017. PMID: 28294537 Review.
Cited by
-
Novel brd4 inhibitors with a unique scaffold exhibit antitumor effects.Oncol Lett. 2021 Jun;21(6):473. doi: 10.3892/ol.2021.12734. Epub 2021 Apr 14. Oncol Lett. 2021. PMID: 33907583 Free PMC article.
-
Methylation as a key regulator of Tau aggregation and neuronal health in Alzheimer's disease.Cell Commun Signal. 2021 May 7;19(1):51. doi: 10.1186/s12964-021-00732-z. Cell Commun Signal. 2021. PMID: 33962636 Free PMC article. Review.
-
Reactivation of latent HIV by histone deacetylase inhibitors.Trends Microbiol. 2013 Jun;21(6):277-85. doi: 10.1016/j.tim.2013.02.005. Epub 2013 Mar 18. Trends Microbiol. 2013. PMID: 23517573 Free PMC article. Review.
-
Targeting Histone Modifications in Bone and Lung Metastatic Cancers.Curr Osteoporos Rep. 2021 Jun;19(3):230-246. doi: 10.1007/s11914-021-00670-2. Epub 2021 Mar 15. Curr Osteoporos Rep. 2021. PMID: 33721181 Free PMC article. Review.
-
Cortactin tyrosine phosphorylation promotes its deacetylation and inhibits cell spreading.PLoS One. 2012;7(3):e33662. doi: 10.1371/journal.pone.0033662. Epub 2012 Mar 30. PLoS One. 2012. PMID: 22479425 Free PMC article.
References
-
- Allis CD, Berger SL, Cote J, Dent S, Jenuwien T, Kouzarides T, Pillus L, Reinberg D, Shi Y, Shiekhattar R, et al. New nomenclature for chromatin-modifying enzymes. Cell. 2007;131:633–636. - PubMed
-
- Appella E, Anderson CW. Signaling to p53: breaking the posttranslational modification code. Pathol Biol (Paris) 2000;48:227–245. - PubMed
-
- Bali P, Pranpat M, Bradner J, Balasis M, Fiskus W, Guo F, Rocha K, Kumaraswamy S, Boyapalle S, Atadja P, et al. Inhibition of histone deacetylase 6 acetylates and disrupts the chaperone function of heat shock protein 90: a novel basis for antileukemia activity of histone deacetylase inhibitors. J Biol Chem. 2005;280:26729–26734. - PubMed
-
- Berger SL. The complex language of chromatin regulation during transcription. Nature. 2007;447:407–412. - PubMed
-
- Bhaumik SR, Smith E, Shilatifard A. Covalent modifications of histones during development and disease pathogenesis. Nat Struct Mol Biol. 2007;14:1008–1016. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
