

Global sequencing of proteolytic cleavage sites in apoptosis by specific labeling of protein N termini
- PMID: 18722006
- PMCID: PMC2566540
- DOI: 10.1016/j.cell.2008.08.012
Global sequencing of proteolytic cleavage sites in apoptosis by specific labeling of protein N termini
Abstract
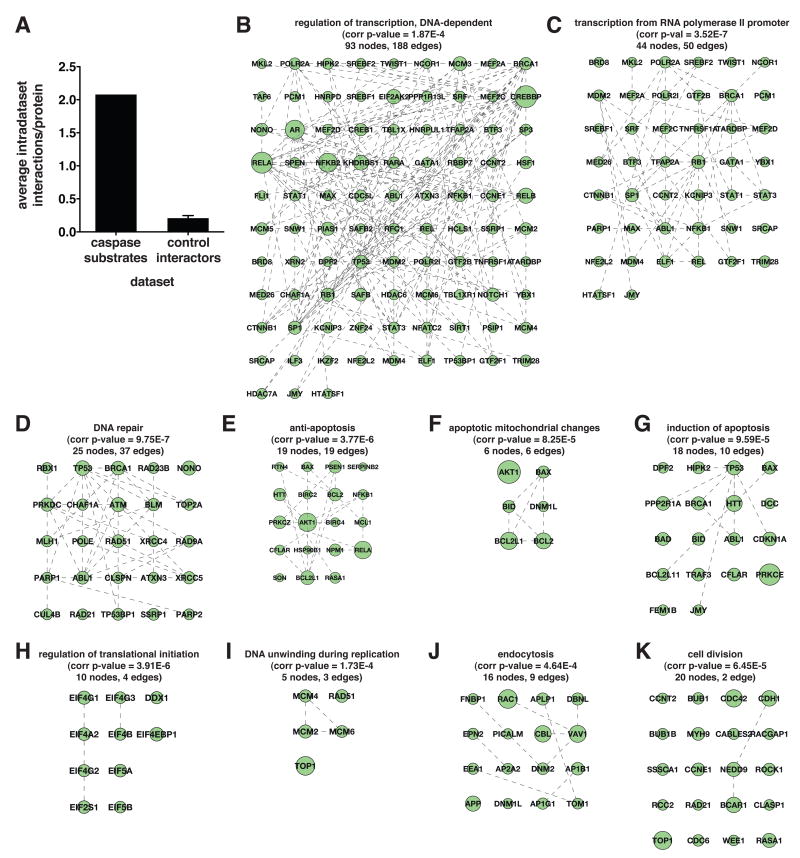
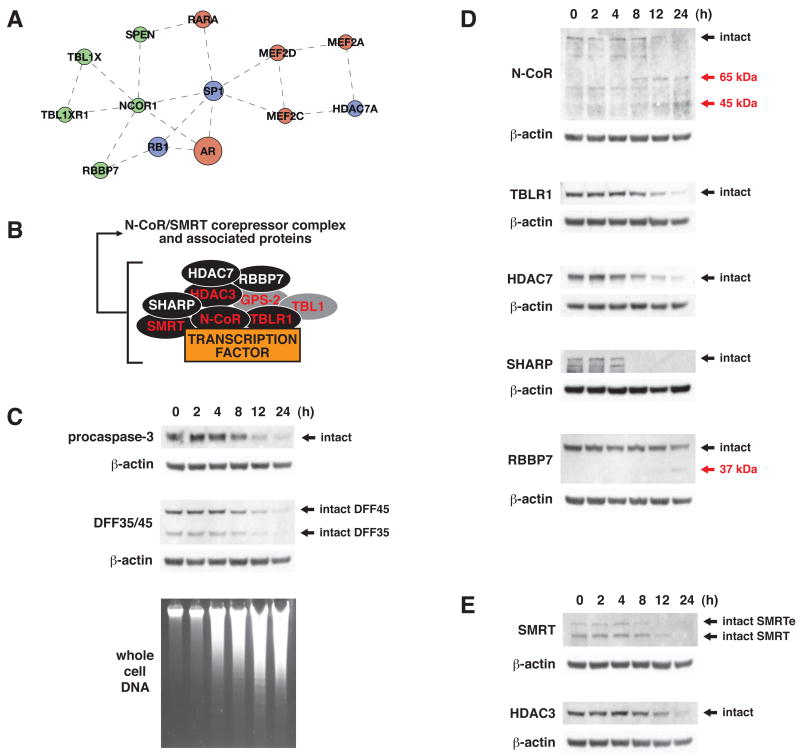
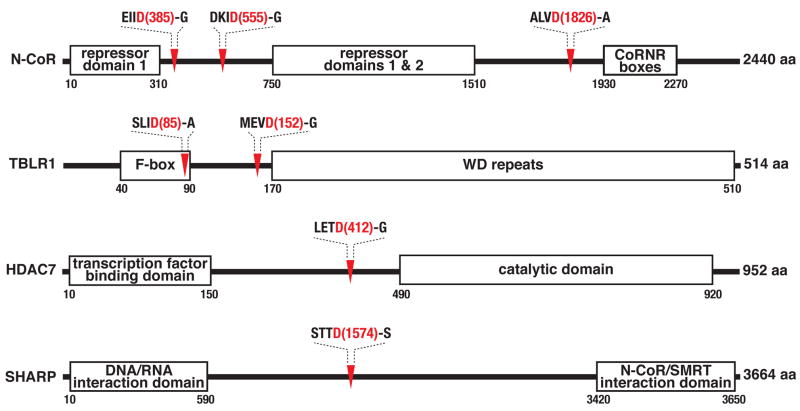
The nearly 600 proteases in the human genome regulate a diversity of biological processes, including programmed cell death. Comprehensive characterization of protease signaling in complex biological samples is limited by available proteomic methods. We have developed a general approach for global identification of proteolytic cleavage sites using an engineered enzyme to selectively biotinylate free protein N termini for positive enrichment of corresponding N-terminal peptides. Using this method to study apoptosis, we have sequenced 333 caspase-like cleavage sites distributed among 292 protein substrates. These sites are generally not predicted by in vitro caspase substrate specificity but can be used to predict other physiological caspase cleavage sites. Structural bioinformatic studies show that caspase cleavage sites often appear in surface-accessible loops and even occasionally in helical regions. Strikingly, we also find that a disproportionate number of caspase substrates physically interact, suggesting that these dimeric proteases target protein complexes and networks to elicit apoptosis.
Figures
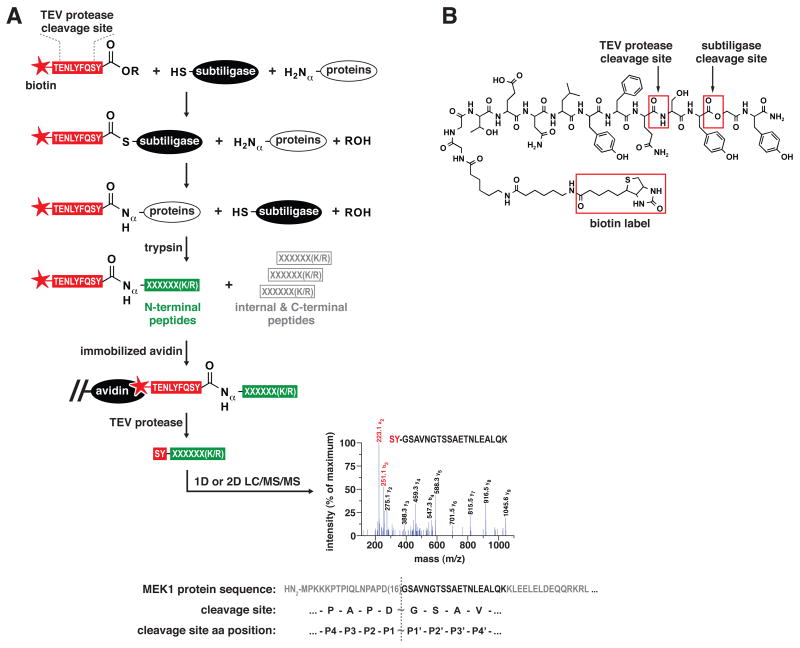
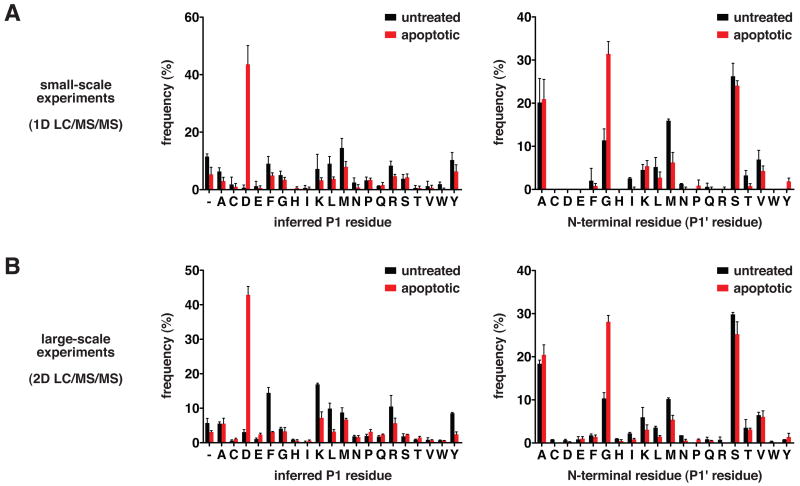
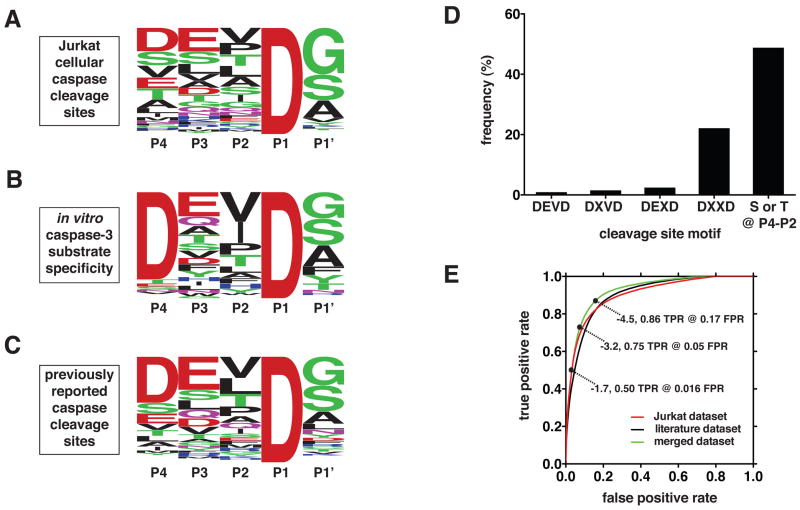
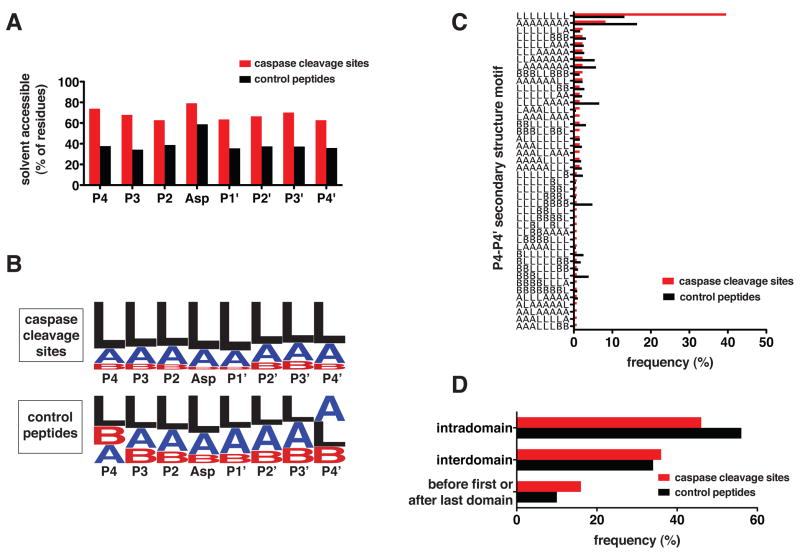
Comment in
-
Caspase cleavage is not for everyone.Cell. 2008 Sep 5;134(5):720-1. doi: 10.1016/j.cell.2008.08.019. Cell. 2008. PMID: 18775303
Similar articles
-
Caspase-specific and nonspecific in vivo protein processing during Fas-induced apoptosis.Nat Methods. 2005 Oct;2(10):771-7. doi: 10.1038/nmeth792. Nat Methods. 2005. PMID: 16179924
-
Complementary proteomic tools for the dissection of apoptotic proteolysis events.J Proteome Res. 2012 May 4;11(5):2947-54. doi: 10.1021/pr300035k. Epub 2012 Mar 29. J Proteome Res. 2012. PMID: 22432722
-
Caspase-8 can be activated by interchain proteolysis without receptor-triggered dimerization during drug-induced apoptosis.J Biol Chem. 2005 Feb 18;280(7):5267-73. doi: 10.1074/jbc.M408585200. Epub 2004 Dec 15. J Biol Chem. 2005. PMID: 15611097
-
Caspases and their substrates.Cell Death Differ. 2017 Aug;24(8):1380-1389. doi: 10.1038/cdd.2017.44. Epub 2017 May 12. Cell Death Differ. 2017. PMID: 28498362 Free PMC article. Review.
-
Caspase substrates: easily caught in deep waters?Trends Biotechnol. 2009 Dec;27(12):680-8. doi: 10.1016/j.tibtech.2009.09.007. Epub 2009 Oct 29. Trends Biotechnol. 2009. PMID: 19879007 Review.
Cited by
-
Engineered cellular gene-replacement platform for selective and inducible proteolytic profiling.Proc Natl Acad Sci U S A. 2015 Jul 7;112(27):8344-9. doi: 10.1073/pnas.1504141112. Epub 2015 Jun 23. Proc Natl Acad Sci U S A. 2015. PMID: 26106156 Free PMC article.
-
Thinking big with small molecules.J Cell Biol. 2015 Apr 13;209(1):7-9. doi: 10.1083/jcb.201501084. J Cell Biol. 2015. PMID: 25869661 Free PMC article.
-
Phage display and structural studies reveal plasticity in substrate specificity of caspase-3a from zebrafish.Protein Sci. 2016 Nov;25(11):2076-2088. doi: 10.1002/pro.3032. Epub 2016 Sep 14. Protein Sci. 2016. PMID: 27577093 Free PMC article.
-
Caspase-dependent Proteolysis of Human Ribonucleotide Reductase Small Subunits R2 and p53R2 during Apoptosis.J Biol Chem. 2015 May 29;290(22):14077-90. doi: 10.1074/jbc.M115.649640. Epub 2015 Apr 15. J Biol Chem. 2015. PMID: 25878246 Free PMC article.
-
The DegraBase: a database of proteolysis in healthy and apoptotic human cells.Mol Cell Proteomics. 2013 Mar;12(3):813-24. doi: 10.1074/mcp.O112.024372. Epub 2012 Dec 20. Mol Cell Proteomics. 2013. PMID: 23264352 Free PMC article.
References
-
- Abrahmsén L, Tom J, Burnier J, Butcher KA, Kossiakoff A, Wells JA. Engineering subtilisin and its substrates for efficient ligation of peptide bonds in aqueous solution. Biochemistry. 1991;30:4151–4159. - PubMed
-
- Berman HM, Battistuz T, Bhat TN, Bluhm WF, Bourne PE, Burkhardt K, Feng Z, Gilliland GL, Iype L, Jain S, et al. The Protein Data Bank. Acta Crystallogr D Biol Crystallogr. 2002;58:899–907. - PubMed
-
- Bolden JE, Peart MJ, Johnstone RW. Anticancer activities of histone deacetylase inhibitors. Nature reviews Drug discovery. 2006;5:769–784. - PubMed
-
- Braisted AC, Judice JK, Wells JA. Synthesis of proteins by subtiligase. Methods Enzymol. 1997;289:298–313. - PubMed
-
- Brown JL, Roberts WK. Evidence that approximately eighty per cent of the soluble proteins from Ehrlich ascites cells are Nalpha-acetylated. J Biol Chem. 1976;251:1009–1014. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F32 GM074458-01A1/GM/NIGMS NIH HHS/United States
- F32 GM074458-03/GM/NIGMS NIH HHS/United States
- R01 GM081051/GM/NIGMS NIH HHS/United States
- R01 GM54762/GM/NIGMS NIH HHS/United States
- P01 GM071790/GM/NIGMS NIH HHS/United States
- U54 GM074945/GM/NIGMS NIH HHS/United States
- F32 GM074458/GM/NIGMS NIH HHS/United States
- P41 RR001614-25/RR/NCRR NIH HHS/United States
- F32 GM074458-02/GM/NIGMS NIH HHS/United States
- U54 GM074945-03S1/GM/NIGMS NIH HHS/United States
- R01 GM081051-02/GM/NIGMS NIH HHS/United States
- R01 GM054762-08/GM/NIGMS NIH HHS/United States
- R01 GM081051-01/GM/NIGMS NIH HHS/United States
- P01 GM71790/GM/NIGMS NIH HHS/United States
- NCRR 01614/PHS HHS/United States
- R01 GM054762/GM/NIGMS NIH HHS/United States
- P01 GM071790-04/GM/NIGMS NIH HHS/United States
- P41 RR001614/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
