

Actin disassembly by cofilin, coronin, and Aip1 occurs in bursts and is inhibited by barbed-end cappers
- PMID: 18663144
- PMCID: PMC2483518
- DOI: 10.1083/jcb.200801027
Actin disassembly by cofilin, coronin, and Aip1 occurs in bursts and is inhibited by barbed-end cappers
Abstract
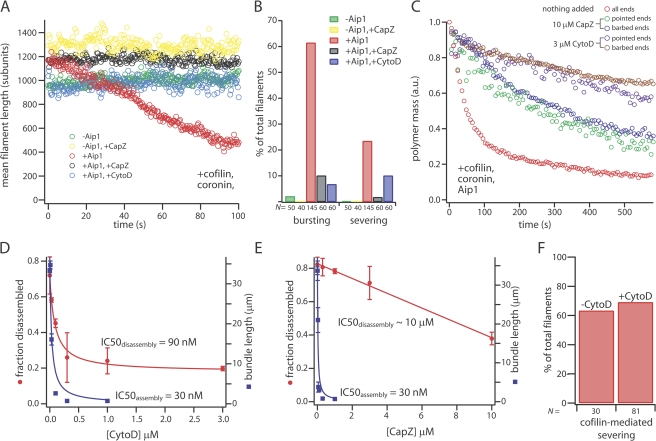
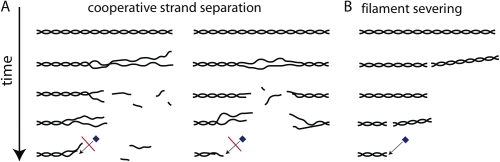
Turnover of actin filaments in cells requires rapid actin disassembly in a cytoplasmic environment that thermodynamically favors assembly because of high concentrations of polymerizable monomers. We here image the disassembly of single actin filaments by cofilin, coronin, and actin-interacting protein 1, a purified protein system that reconstitutes rapid, monomer-insensitive disassembly (Brieher, W.M., H.Y. Kueh, B.A. Ballif, and T.J. Mitchison. 2006. J. Cell Biol. 175:315-324). In this three-component system, filaments disassemble in abrupt bursts that initiate preferentially, but not exclusively, from both filament ends. Bursting disassembly generates unstable reaction intermediates with lowered affinity for CapZ at barbed ends. CapZ and cytochalasin D (CytoD), a barbed-end capping drug, strongly inhibit bursting disassembly. CytoD also inhibits actin disassembly in mammalian cells, whereas latrunculin B, a monomer sequestering drug, does not. We propose that bursts of disassembly arise from cooperative separation of the two filament strands near an end. The differential effects of drugs in cells argue for physiological relevance of this new disassembly pathway and potentially explain discordant results previously found with these drugs.
Figures






Similar articles
-
Loss of Aip1 reveals a role in maintaining the actin monomer pool and an in vivo oligomer assembly pathway.J Cell Biol. 2010 Mar 22;188(6):769-77. doi: 10.1083/jcb.200909176. Epub 2010 Mar 15. J Cell Biol. 2010. PMID: 20231387 Free PMC article.
-
Catastrophic actin filament bursting by cofilin, Aip1, and coronin.J Biol Chem. 2020 Sep 18;295(38):13299-13313. doi: 10.1074/jbc.RA120.015018. Epub 2020 Jul 28. J Biol Chem. 2020. PMID: 32723865 Free PMC article.
-
Microscopic evidence that actin-interacting protein 1 actively disassembles actin-depolymerizing factor/Cofilin-bound actin filaments.J Biol Chem. 2004 Apr 2;279(14):14207-12. doi: 10.1074/jbc.M313418200. Epub 2004 Jan 23. J Biol Chem. 2004. PMID: 14742433
-
Control of actin assembly and disassembly at filament ends.Curr Opin Cell Biol. 2000 Feb;12(1):97-103. doi: 10.1016/s0955-0674(99)00062-9. Curr Opin Cell Biol. 2000. PMID: 10679358 Review.
-
Coronin: the double-edged sword of actin dynamics.Subcell Biochem. 2008;48:72-87. doi: 10.1007/978-0-387-09595-0_7. Subcell Biochem. 2008. PMID: 18925372 Review.
Cited by
-
Srv2/cyclase-associated protein forms hexameric shurikens that directly catalyze actin filament severing by cofilin.Mol Biol Cell. 2013 Jan;24(1):31-41. doi: 10.1091/mbc.E12-08-0589. Epub 2012 Nov 7. Mol Biol Cell. 2013. PMID: 23135996 Free PMC article.
-
Plasma membrane calcium ATPase activity is regulated by actin oligomers through direct interaction.J Biol Chem. 2013 Aug 9;288(32):23380-93. doi: 10.1074/jbc.M113.470542. Epub 2013 Jun 26. J Biol Chem. 2013. PMID: 23803603 Free PMC article.
-
Evolutionary selection of new breast cancer cell-targeting peptides and phages with the cell-targeting peptides fully displayed on the major coat and their effects on actin dynamics during cell internalization.Mol Pharm. 2010 Oct 4;7(5):1629-42. doi: 10.1021/mp100052y. Epub 2010 Aug 25. Mol Pharm. 2010. PMID: 20735141 Free PMC article.
-
Functional surfaces on the actin-binding protein coronin revealed by systematic mutagenesis.J Biol Chem. 2010 Nov 5;285(45):34899-908. doi: 10.1074/jbc.M110.171496. Epub 2010 Sep 2. J Biol Chem. 2010. PMID: 20813846 Free PMC article.
-
The QKI-6 RNA binding protein regulates actin-interacting protein-1 mRNA stability during oligodendrocyte differentiation.Mol Biol Cell. 2010 Sep 1;21(17):3029-40. doi: 10.1091/mbc.E10-04-0305. Epub 2010 Jul 14. Mol Biol Cell. 2010. PMID: 20631256 Free PMC article.
References
-
- Amberg, D.C., E. Basart, and D. Botstein. 1995. Defining protein interactions with yeast actin in vivo. Nat. Struct. Biol. 2:28–35. - PubMed
-
- Andrianantoandro, E., and T.D. Pollard. 2006. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol. Cell. 24:13–23. - PubMed
-
- Balcer, H.I., A.L. Goodman, A.A. Rodal, E. Smith, J. Kugler, J.E. Heuser, and B.L. Goode. 2003. Coordinated regulation of actin filament turnover by a high-molecular-weight Srv2/CAP complex, cofilin, profilin, and Aip1. Curr. Biol. 13:2159–2169. - PubMed
-
- Blanchoin, L., and T.D. Pollard. 1999. Mechanism of interaction of Acanthamoeba actophorin (ADF/Cofilin) with actin filaments. J. Biol. Chem. 274:15538–15546. - PubMed
-
- Bobkov, A.A., A. Muhlrad, A. Shvetsov, S. Benchaar, D. Scoville, S.C. Almo, and E. Reisler. 2004. Cofilin (ADF) affects lateral contacts in F-actin. J. Mol. Biol. 337:93–104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
