

Systems biology of vascular endothelial growth factors
- PMID: 18608994
- PMCID: PMC2722038
- DOI: 10.1080/10739680802095964
Systems biology of vascular endothelial growth factors
Abstract
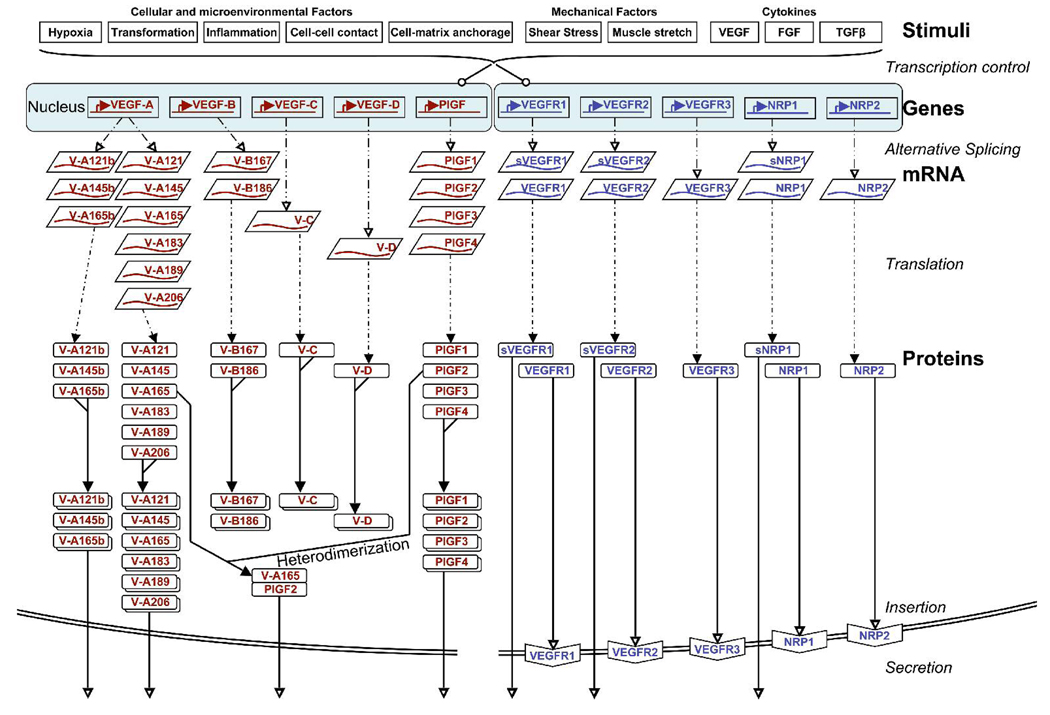
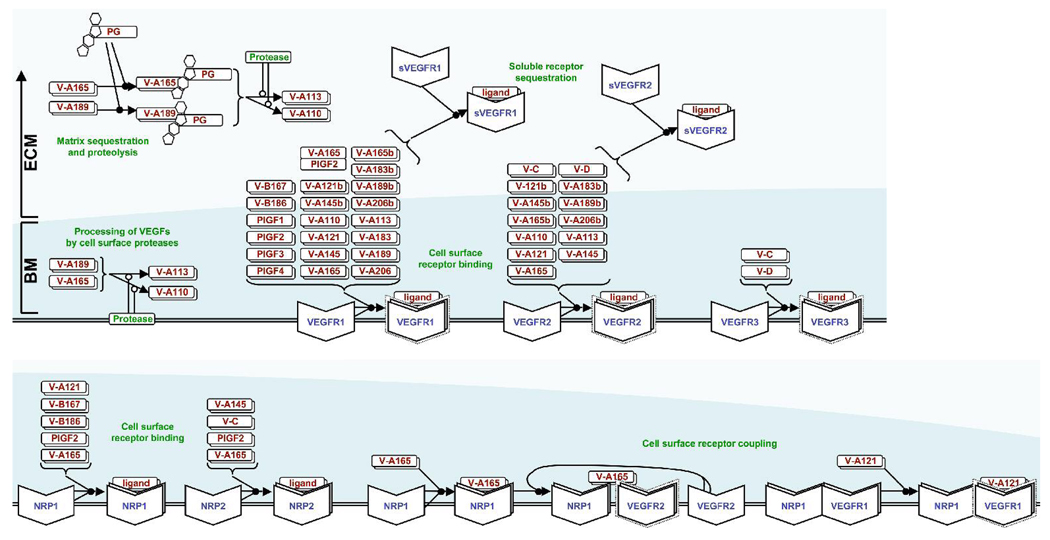
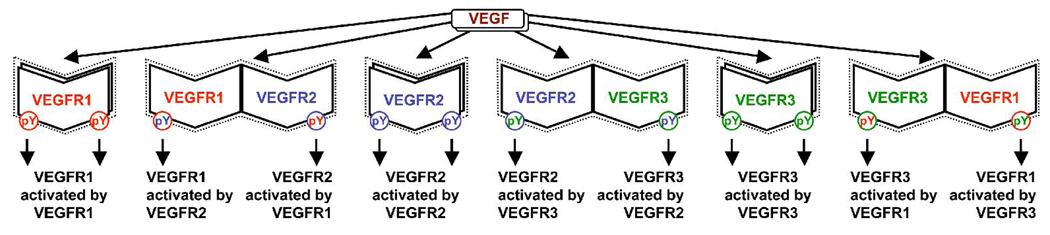
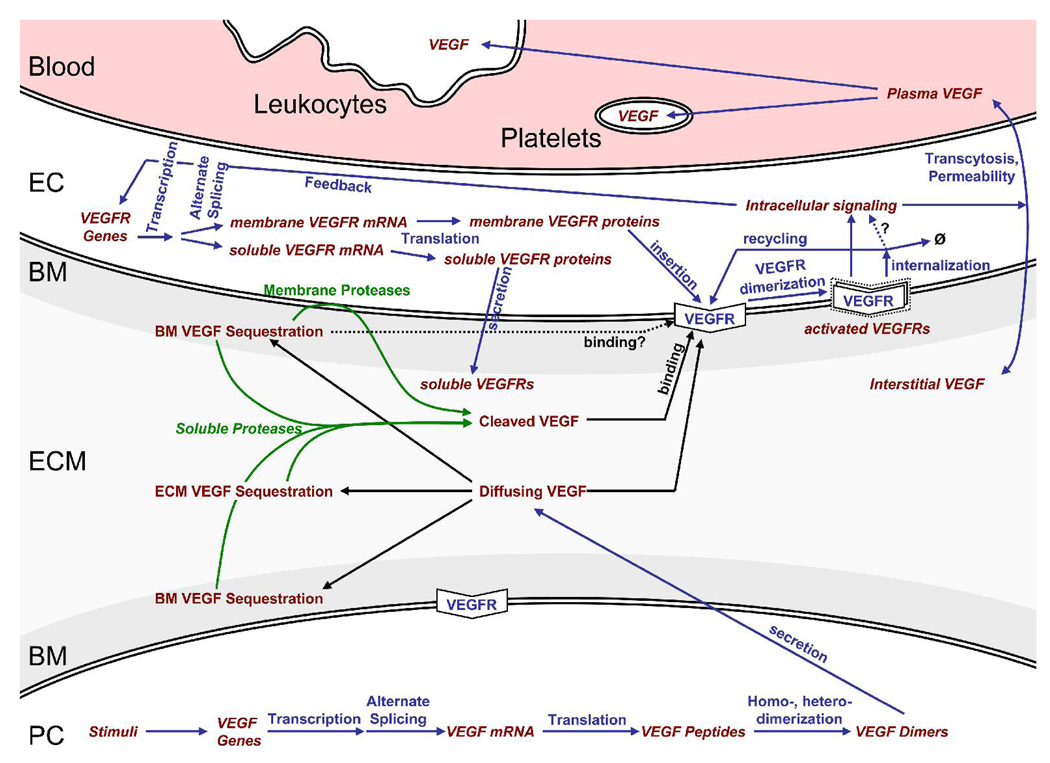
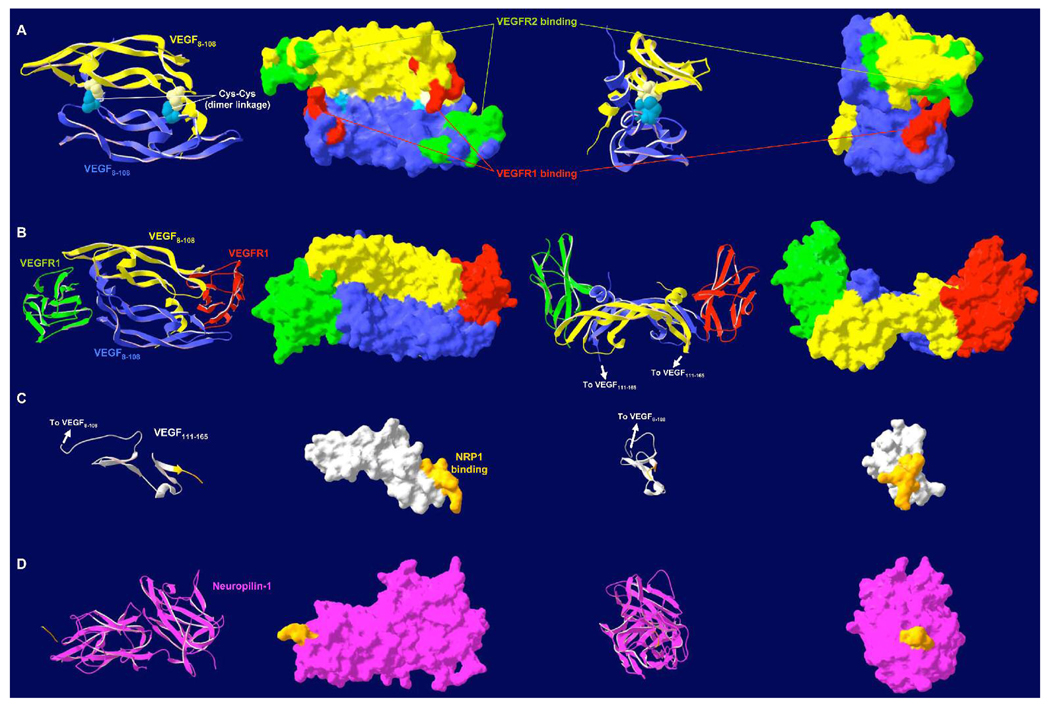
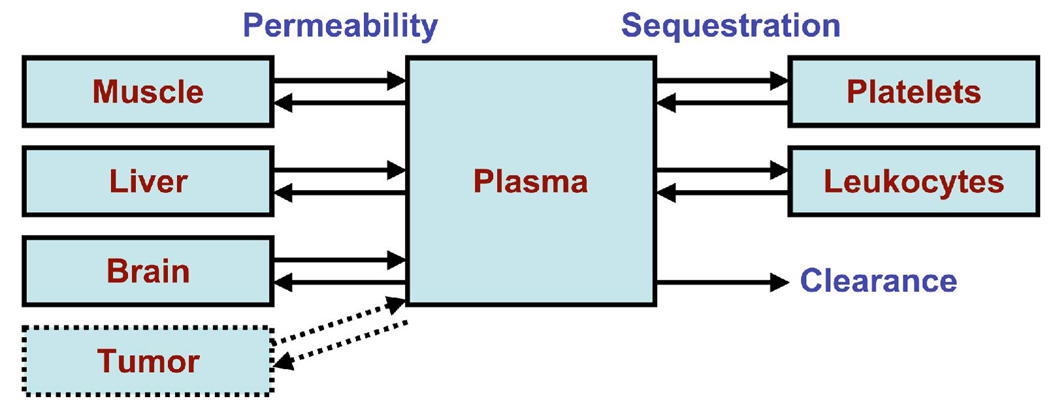
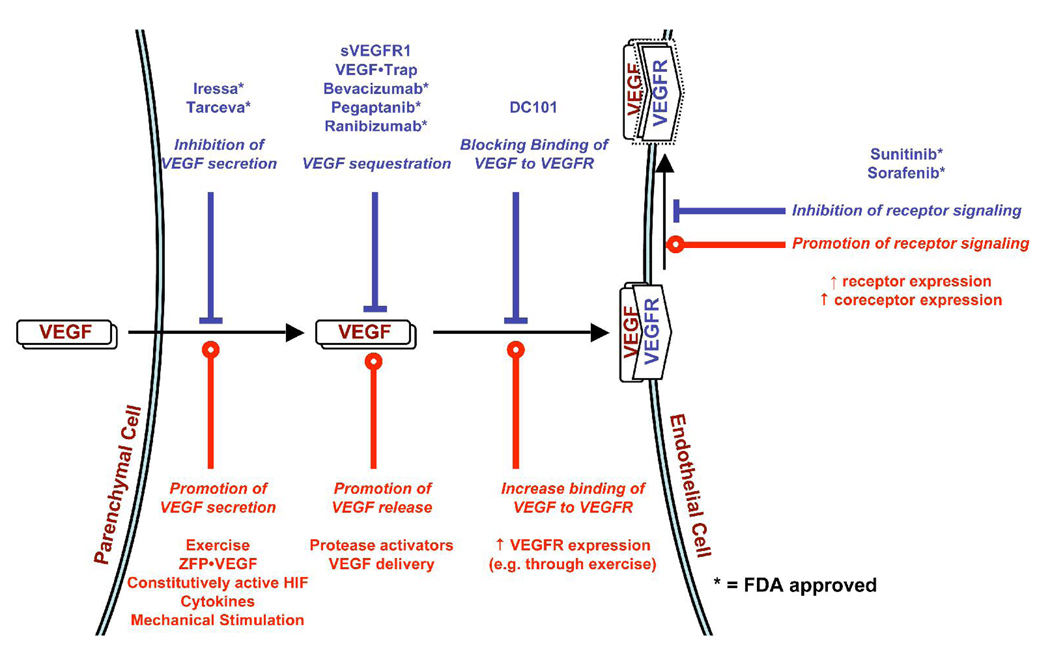
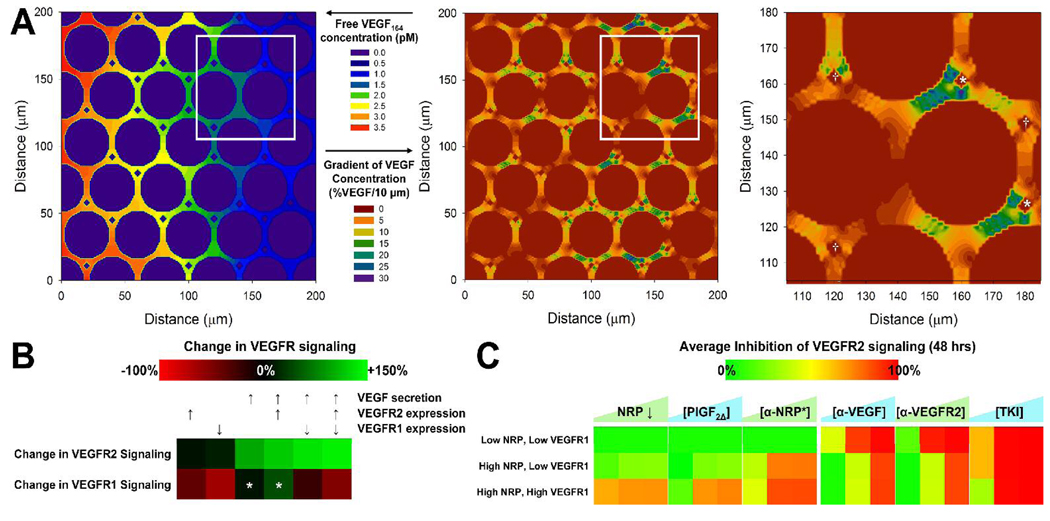
Several cytokine families have roles in the development, maintenance, and remodeling of the microcirculation. Of these, the vascular endothelial growth factor (VEGF) family is one of the best studied and one of the most complex. Five VEGF ligand genes and five cell-surface receptor genes are known in the human, and each of these may be transcribed as multiple splice isoforms to generate an extensive family of proteins, many of which are subject to further proteolytic processing. Using the VEGF family as an example, we describe the current knowledge of growth-factor expression, processing, and transport in vivo. Experimental studies and computational simulations are being used to measure and predict the activity of these molecules, and we describe avenues of research that seek to fill the remaining gaps in our understanding of VEGF family behavior.
Figures
Similar articles
-
Modeling of growth factor-receptor systems from molecular-level protein interaction networks to whole-body compartment models.Methods Enzymol. 2009;467:461-497. doi: 10.1016/S0076-6879(09)67018-X. Methods Enzymol. 2009. PMID: 19897104 Free PMC article.
-
Role of the vascular endothelial growth factor isoforms in retinal angiogenesis and DiGeorge syndrome.Verh K Acad Geneeskd Belg. 2005;67(4):229-76. Verh K Acad Geneeskd Belg. 2005. PMID: 16334858 Review.
-
Molecular and functional diversity of vascular endothelial growth factors.Mol Divers. 2006 Nov;10(4):515-27. doi: 10.1007/s11030-006-9027-3. Mol Divers. 2006. PMID: 16972015 Review.
-
Mathematical Modeling of Cellular Cross-Talk Between Endothelial and Tumor Cells Highlights Counterintuitive Effects of VEGF-Targeted Therapies.Bull Math Biol. 2018 May;80(5):971-1016. doi: 10.1007/s11538-017-0273-6. Epub 2017 Apr 24. Bull Math Biol. 2018. PMID: 28439752
-
[Tumor angiogenesis and its inhibitors].Nihon Rinsho. 2012 Nov;70 Suppl 8:149-53. Nihon Rinsho. 2012. PMID: 23513829 Japanese. No abstract available.
Cited by
-
Understanding vascular development.Wiley Interdiscip Rev Dev Biol. 2013 May-Jun;2(3):327-46. doi: 10.1002/wdev.91. Epub 2012 Oct 5. Wiley Interdiscip Rev Dev Biol. 2013. PMID: 23799579 Free PMC article. Review.
-
Computational models of VEGF-associated angiogenic processes in cancer.Math Med Biol. 2012 Mar;29(1):85-94. doi: 10.1093/imammb/dqq025. Epub 2011 Jan 25. Math Med Biol. 2012. PMID: 21266494 Free PMC article.
-
Quantification and cell-to-cell variation of vascular endothelial growth factor receptors.Exp Cell Res. 2011 Apr 15;317(7):955-65. doi: 10.1016/j.yexcr.2010.12.014. Epub 2010 Dec 23. Exp Cell Res. 2011. PMID: 21185287 Free PMC article.
-
Retinoblastoma binding protein 2 (RBP2) promotes HIF-1α-VEGF-induced angiogenesis of non-small cell lung cancer via the Akt pathway.PLoS One. 2014 Aug 27;9(8):e106032. doi: 10.1371/journal.pone.0106032. eCollection 2014. PLoS One. 2014. PMID: 25162518 Free PMC article.
-
Pericellular proteolysis in cancer.Genes Dev. 2014 Nov 1;28(21):2331-47. doi: 10.1101/gad.250647.114. Genes Dev. 2014. PMID: 25367033 Free PMC article. Review.
References
-
- Ahmad S, Ahmed A. Elevated placental soluble vascular endothelial growth factor receptor-1 inhibits angiogenesis in preeclampsia. Circ Res. 2004;95:884–891. - PubMed
-
- Alam A, Herault JP, Barron P, Favier B, Fons P, Delesque-Touchard N, Senegas I, Laboudie P, Bonnin J, Cassan C, Savi P, Ruggeri B, Carmeliet P, Bono F, Herbert JM. Heterodimerization with vascular endothelial growth factor receptor-2 (VEGFR-2) is necessary for VEGFR-3 activity. Biochem Biophys Res Commun. 2004;324:909–915. - PubMed
-
- Amaral SL, Linderman JR, Morse MM, Greene AS. Angiogenesis induced by electrical stimulation is mediated by angiotensin II and VEGF. Microcirculation. 2001;8:57–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
