

Trans-cellular migration: cell-cell contacts get intimate
- PMID: 18595683
- PMCID: PMC2811962
- DOI: 10.1016/j.ceb.2008.05.007
Trans-cellular migration: cell-cell contacts get intimate
Abstract
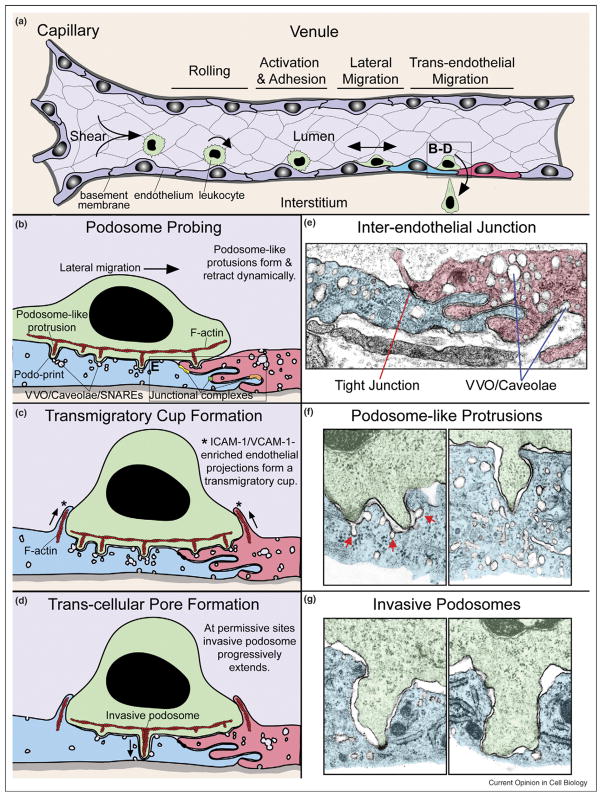
Trans-cellular migration, the movement of one cell directly through another, seems an unlikely, counterintuitive, and even bizarre process. Trans-cellular migration has been reported for nearly half a century in leukocyte transendothelial migration in vivo, but is not well enough accepted to widely feature in textbook accounts of diapedesis. Recently, the first in vitro and additional in vivo observations of trans-cellular diapedesis have been reported. Mechanisms by which this occurs are just beginning to be elucidated and point to podosome-like protrusive activities in leukocytes and specific fusogenic functions in endothelial cells. Emerging evidence for a quantitatively significant contribution of trans-cellular migration to leukocyte trafficking in increasingly diverse settings suggests that this phenomenon represents an important and physiologic cell biological process.
Figures
Similar articles
-
Settings and mechanisms for trans-cellular diapedesis.Front Biosci (Landmark Ed). 2009 Jun 1;14(13):5066-83. doi: 10.2741/3587. Front Biosci (Landmark Ed). 2009. PMID: 19482605 Free PMC article. Review.
-
Lymphocyte transcellular migration occurs through recruitment of endothelial ICAM-1 to caveola- and F-actin-rich domains.Nat Cell Biol. 2006 Feb;8(2):113-23. doi: 10.1038/ncb1356. Epub 2006 Jan 22. Nat Cell Biol. 2006. PMID: 16429128
-
Analysis of flow-based adhesion in vitro.Methods Mol Med. 2007;135:323-31. doi: 10.1007/978-1-59745-401-8_20. Methods Mol Med. 2007. PMID: 17951668
-
Molecular events during leukocyte diapedesis.FEBS J. 2006 Oct;273(19):4399-407. doi: 10.1111/j.1742-4658.2006.05439.x. Epub 2006 Sep 11. FEBS J. 2006. PMID: 16965541 Review.
-
Simultaneous tether extraction from endothelial cells and leukocytes: observation, mechanics, and significance.Biophys J. 2007 Dec 1;93(11):4041-52. doi: 10.1529/biophysj.107.109298. Epub 2007 Aug 17. Biophys J. 2007. PMID: 17704170 Free PMC article.
Cited by
-
Collagen inhibitory peptide R1R2 mediates vascular remodeling by decreasing inflammation and smooth muscle cell activation.PLoS One. 2015 Feb 12;10(2):e0117356. doi: 10.1371/journal.pone.0117356. eCollection 2015. PLoS One. 2015. PMID: 25675397 Free PMC article.
-
Endothelial junction regulation: a prerequisite for leukocytes crossing the vessel wall.J Innate Immun. 2013;5(4):324-35. doi: 10.1159/000348828. Epub 2013 Apr 3. J Innate Immun. 2013. PMID: 23571667 Free PMC article. Review.
-
Myosin light chain kinase in microvascular endothelial barrier function.Cardiovasc Res. 2010 Jul 15;87(2):272-80. doi: 10.1093/cvr/cvq144. Epub 2010 May 17. Cardiovasc Res. 2010. PMID: 20479130 Free PMC article. Review.
-
Septin and actin contributions to endothelial cell-cell junctions and monolayer integrity.Cytoskeleton (Hoboken). 2023 Jul-Aug;80(7-8):228-241. doi: 10.1002/cm.21732. Epub 2022 Oct 17. Cytoskeleton (Hoboken). 2023. PMID: 36205643 Free PMC article. Review.
-
Mechanisms of leukocyte transendothelial migration.Annu Rev Pathol. 2011;6:323-44. doi: 10.1146/annurev-pathol-011110-130224. Annu Rev Pathol. 2011. PMID: 21073340 Free PMC article. Review.
References
-
- von Andrian UH, Mackay CR. T-cell function and migration. Two sides of the same coin. N Engl J Med. 2000;343:1020–1034. - PubMed
-
- Aird WC. Phenotypic heterogeneity of the endothelium. I. Structure, function, and mechanisms. Circ Res. 2007;100:158–173. - PubMed
-
- Aird WC. Phenotypic heterogeneity of the endothelium. II. Representative vascular beds. Circ Res. 2007;100:174–190. - PubMed
-
- Bazzoni G, Dejana E. Endothelial cell-to-cell junctions: molecular organization and role in vascular homeostasis. Physiol Rev. 2004;84:869–901. - PubMed
-
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multi-step paradigm. Cell. 1994;76:301–314. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
