

SMAD proteins control DROSHA-mediated microRNA maturation
- PMID: 18548003
- PMCID: PMC2653422
- DOI: 10.1038/nature07086
SMAD proteins control DROSHA-mediated microRNA maturation
Abstract
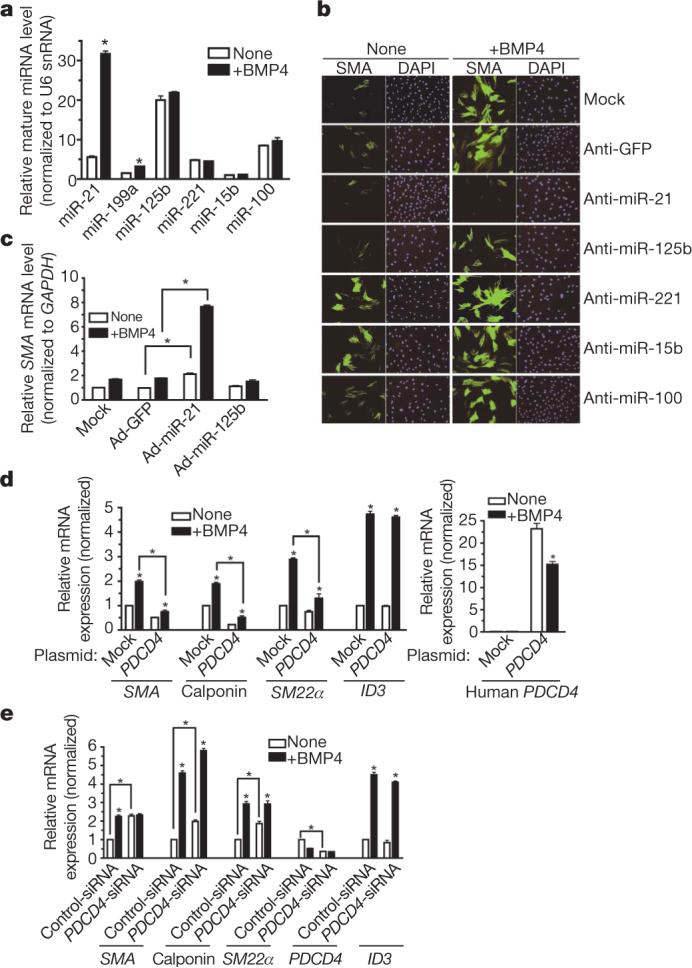
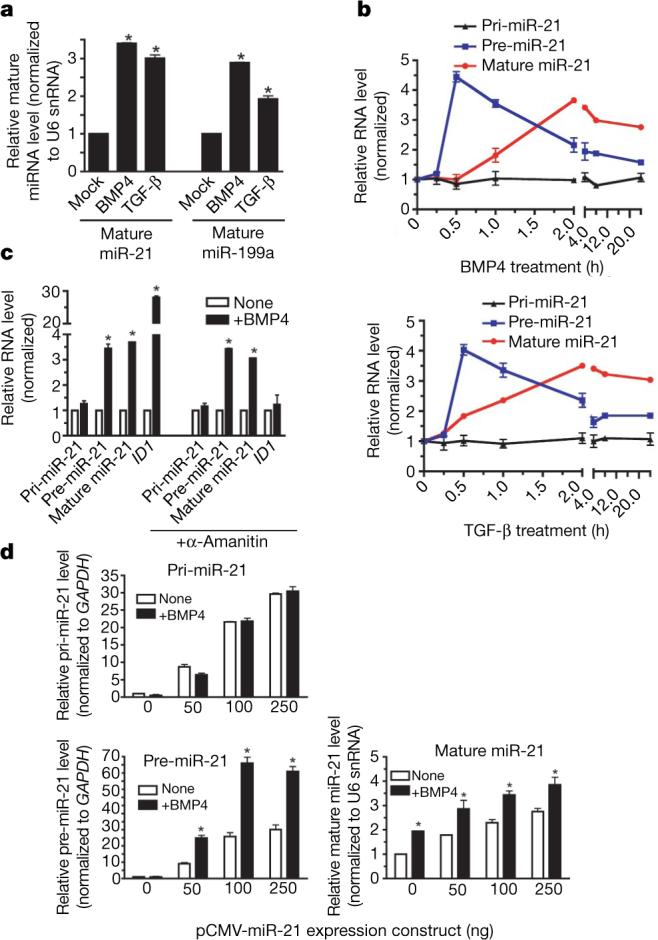
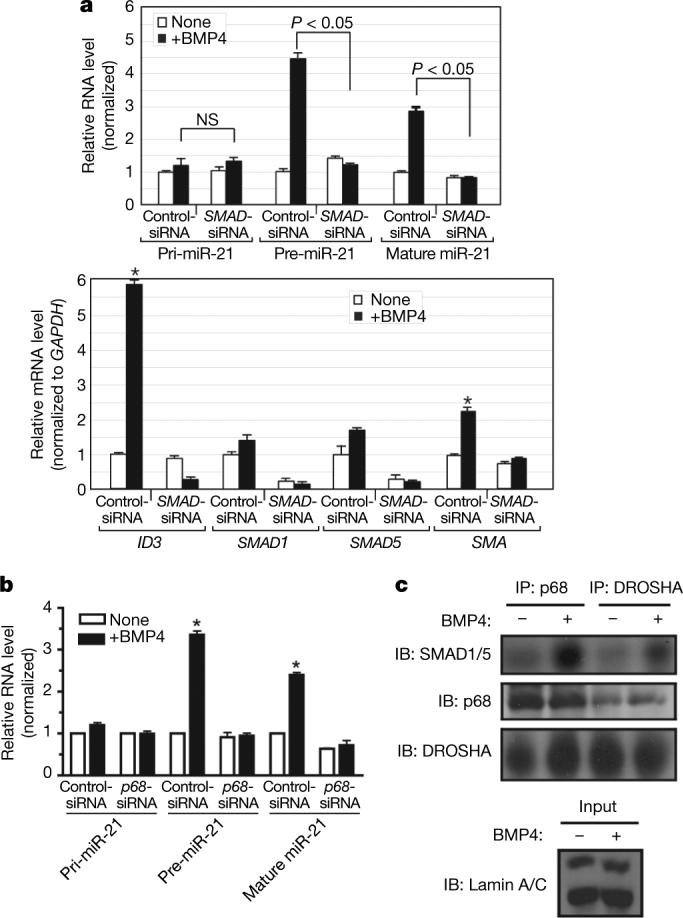
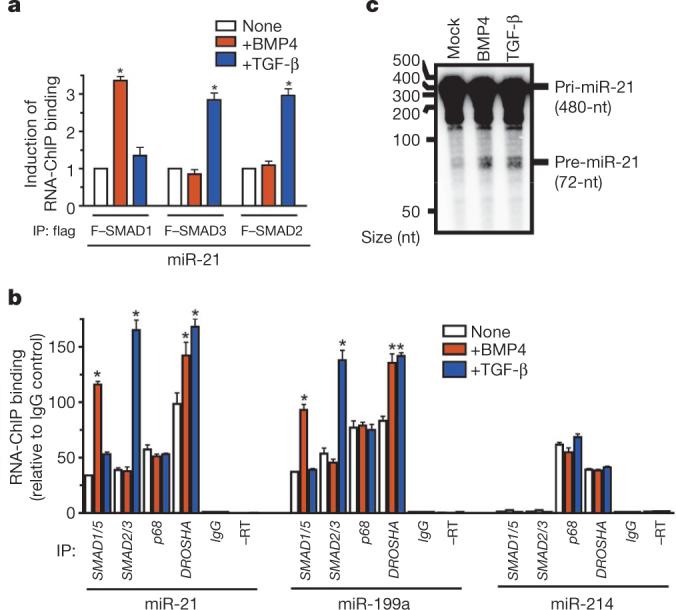
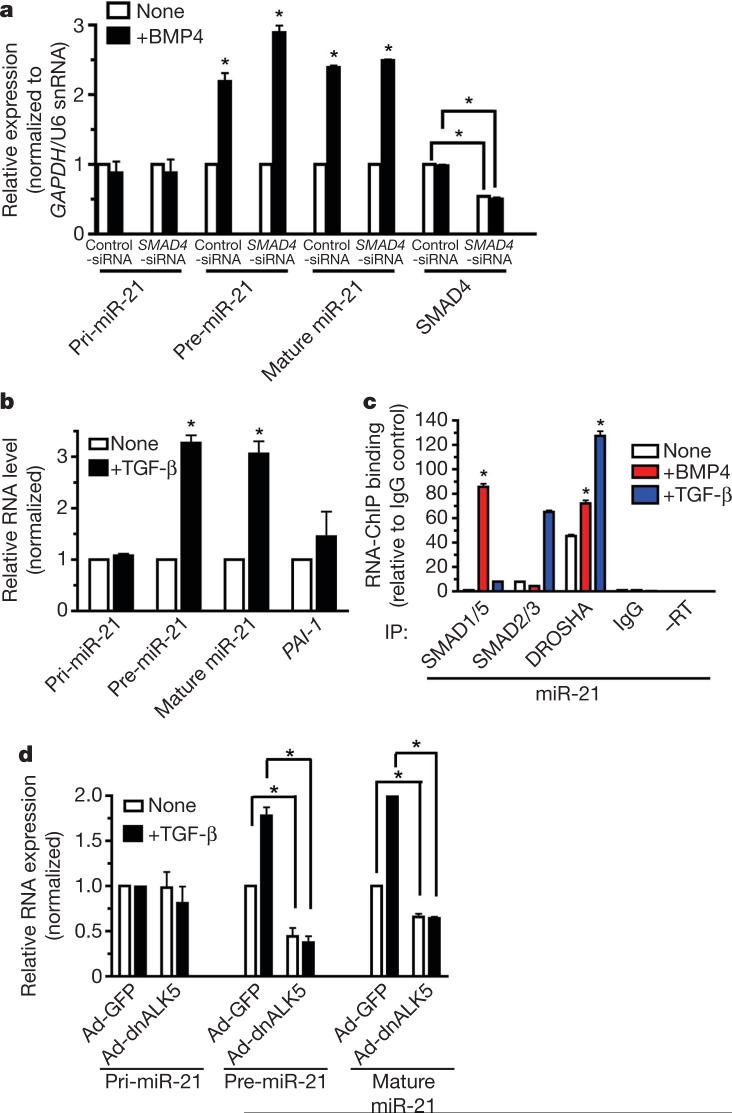
MicroRNAs (miRNAs) are small non-coding RNAs that participate in the spatiotemporal regulation of messenger RNA and protein synthesis. Aberrant miRNA expression leads to developmental abnormalities and diseases, such as cardiovascular disorders and cancer; however, the stimuli and processes regulating miRNA biogenesis are largely unknown. The transforming growth factor beta (TGF-beta) and bone morphogenetic protein (BMP) family of growth factors orchestrates fundamental biological processes in development and in the homeostasis of adult tissues, including the vasculature. Here we show that induction of a contractile phenotype in human vascular smooth muscle cells by TGF-beta and BMPs is mediated by miR-21. miR-21 downregulates PDCD4 (programmed cell death 4), which in turn acts as a negative regulator of smooth muscle contractile genes. Surprisingly, TGF-beta and BMP signalling promotes a rapid increase in expression of mature miR-21 through a post-transcriptional step, promoting the processing of primary transcripts of miR-21 (pri-miR-21) into precursor miR-21 (pre-miR-21) by the DROSHA (also known as RNASEN) complex. TGF-beta- and BMP-specific SMAD signal transducers are recruited to pri-miR-21 in a complex with the RNA helicase p68 (also known as DDX5), a component of the DROSHA microprocessor complex. The shared cofactor SMAD4 is not required for this process. Thus, regulation of miRNA biogenesis by ligand-specific SMAD proteins is critical for control of the vascular smooth muscle cell phenotype and potentially for SMAD4-independent responses mediated by the TGF-beta and BMP signalling pathways.
Figures
Similar articles
-
Smad-mediated regulation of microRNA biosynthesis.FEBS Lett. 2012 Jul 4;586(14):1906-12. doi: 10.1016/j.febslet.2012.01.041. Epub 2012 Jan 28. FEBS Lett. 2012. PMID: 22306316 Free PMC article. Review.
-
Smad proteins bind a conserved RNA sequence to promote microRNA maturation by Drosha.Mol Cell. 2010 Aug 13;39(3):373-84. doi: 10.1016/j.molcel.2010.07.011. Mol Cell. 2010. PMID: 20705240 Free PMC article.
-
down-regulation of Kruppel-like factor-4 (KLF4) by microRNA-143/145 is critical for modulation of vascular smooth muscle cell phenotype by transforming growth factor-beta and bone morphogenetic protein 4.J Biol Chem. 2011 Aug 12;286(32):28097-110. doi: 10.1074/jbc.M111.236950. Epub 2011 Jun 13. J Biol Chem. 2011. PMID: 21673106 Free PMC article.
-
BRCA1 regulates microRNA biogenesis via the DROSHA microprocessor complex.J Cell Biol. 2012 Apr 16;197(2):201-8. doi: 10.1083/jcb.201110008. Epub 2012 Apr 9. J Cell Biol. 2012. PMID: 22492723 Free PMC article.
-
Regulation of miRNA biogenesis as an integrated component of growth factor signaling.Curr Opin Cell Biol. 2013 Apr;25(2):233-40. doi: 10.1016/j.ceb.2012.12.005. Epub 2013 Jan 8. Curr Opin Cell Biol. 2013. PMID: 23312066 Free PMC article. Review.
Cited by
-
Panels of circulating microRNAs as potential diagnostic biomarkers for breast cancer: a systematic review and meta-analysis.Breast Cancer Res Treat. 2022 Nov;196(1):1-15. doi: 10.1007/s10549-022-06728-8. Epub 2022 Sep 9. Breast Cancer Res Treat. 2022. PMID: 36085533 Review.
-
Molecular pathogenesis of pulmonary arterial hypertension.J Clin Invest. 2012 Dec;122(12):4306-13. doi: 10.1172/JCI60658. Epub 2012 Dec 3. J Clin Invest. 2012. PMID: 23202738 Free PMC article. Review.
-
Recombinant adeno-associated virus-mediated inhibition of microRNA-21 protects mice against the lethal schistosome infection by repressing both IL-13 and transforming growth factor beta 1 pathways.Hepatology. 2015 Jun;61(6):2008-2017. doi: 10.1002/hep.27671. Epub 2015 Apr 15. Hepatology. 2015. PMID: 25546547 Free PMC article.
-
TGF-β Signaling from Receptors to Smads.Cold Spring Harb Perspect Biol. 2016 Sep 1;8(9):a022061. doi: 10.1101/cshperspect.a022061. Cold Spring Harb Perspect Biol. 2016. PMID: 27449815 Free PMC article. Review.
-
The harmonies played by TGF-β in stem cell biology.Cell Stem Cell. 2012 Dec 7;11(6):751-64. doi: 10.1016/j.stem.2012.11.001. Cell Stem Cell. 2012. PMID: 23217421 Free PMC article. Review.
References
-
- ten Dijke P, Arthur HM. Extracellular control of TGFβ signalling in vascular development and disease. Nature Rev. Mol. Cell Biol. 2007;8:857–868. - PubMed
-
- Morrell NW. Pulmonary hypertension due to BMPR2 mutation: a new paradigm for tissue remodeling? Proc. Am. Thorac. Soc. 2006;3:680–686. - PubMed
-
- Owens GK. Regulation of differentiation of vascular smooth muscle cells. Physiol. Rev. 1995;75:487–517. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
