

Pyruvate dehydrogenase complex activity controls metabolic and malignant phenotype in cancer cells
- PMID: 18541534
- PMCID: PMC2504897
- DOI: 10.1074/jbc.M801765200
Pyruvate dehydrogenase complex activity controls metabolic and malignant phenotype in cancer cells
Abstract
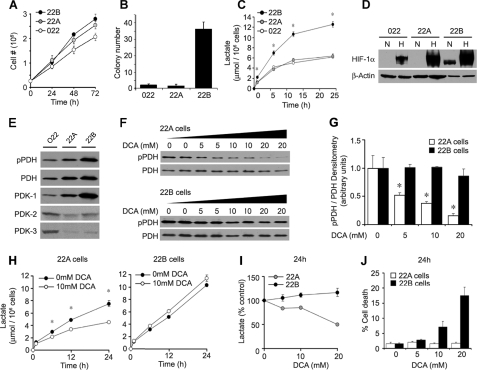
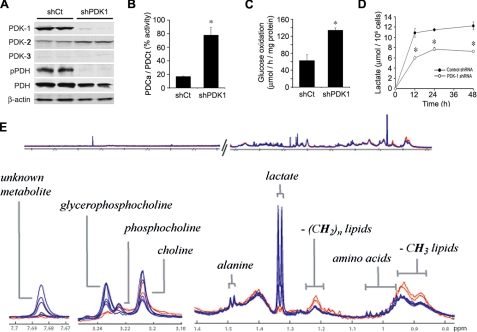
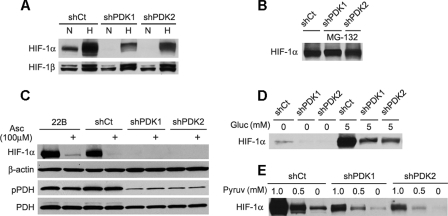
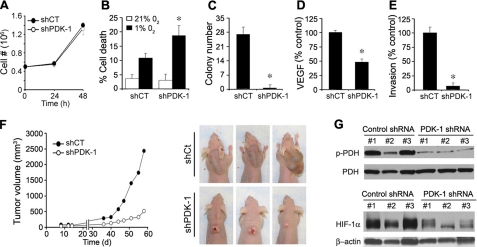
High lactate generation and low glucose oxidation, despite normal oxygen conditions, are commonly seen in cancer cells and tumors. Historically known as the Warburg effect, this altered metabolic phenotype has long been correlated with malignant progression and poor clinical outcome. However, the mechanistic relationship between altered glucose metabolism and malignancy remains poorly understood. Here we show that inhibition of pyruvate dehydrogenase complex (PDC) activity contributes to the Warburg metabolic and malignant phenotype in human head and neck squamous cell carcinoma. PDC inhibition occurs via enhanced expression of pyruvate dehydrogenase kinase-1 (PDK-1), which results in inhibitory phosphorylation of the pyruvate dehydrogenase alpha (PDHalpha) subunit. We also demonstrate that PDC inhibition in cancer cells is associated with normoxic stabilization of the malignancy-promoting transcription factor hypoxia-inducible factor-1alpha (HIF-1alpha) by glycolytic metabolites. Knockdown of PDK-1 via short hairpin RNA lowers PDHalpha phosphorylation, restores PDC activity, reverts the Warburg metabolic phenotype, decreases normoxic HIF-1alpha expression, lowers hypoxic cell survival, decreases invasiveness, and inhibits tumor growth. PDK-1 is an HIF-1-regulated gene, and these data suggest that the buildup of glycolytic metabolites, resulting from high PDK-1 expression, may in turn promote HIF-1 activation, thus sustaining a feed-forward loop for malignant progression. In addition to providing anabolic support for cancer cells, altered fuel metabolism thus supports a malignant phenotype. Correction of metabolic abnormalities offers unique opportunities for cancer treatment and may potentially synergize with other cancer therapies.
Figures
Similar articles
-
Therapeutic Targeting of the Pyruvate Dehydrogenase Complex/Pyruvate Dehydrogenase Kinase (PDC/PDK) Axis in Cancer.J Natl Cancer Inst. 2017 Nov 1;109(11). doi: 10.1093/jnci/djx071. J Natl Cancer Inst. 2017. PMID: 29059435 Review.
-
Targeting pyruvate dehydrogenase kinase signaling in the development of effective cancer therapy.Biochim Biophys Acta Rev Cancer. 2021 Aug;1876(1):188568. doi: 10.1016/j.bbcan.2021.188568. Epub 2021 May 21. Biochim Biophys Acta Rev Cancer. 2021. PMID: 34023419 Review.
-
Nodal regulates energy metabolism in glioma cells by inducing expression of hypoxia-inducible factor 1α.Neuro Oncol. 2013 Oct;15(10):1330-41. doi: 10.1093/neuonc/not086. Epub 2013 Aug 1. Neuro Oncol. 2013. PMID: 23911596 Free PMC article.
-
Anti-Warburg Effect of Melatonin: A Proposed Mechanism to Explain its Inhibition of Multiple Diseases.Int J Mol Sci. 2021 Jan 14;22(2):764. doi: 10.3390/ijms22020764. Int J Mol Sci. 2021. PMID: 33466614 Free PMC article. Review.
-
Expression of pyruvate dehydrogenase kinase-1 in gastric cancer as a potential therapeutic target.Int J Oncol. 2013 Jan;42(1):44-54. doi: 10.3892/ijo.2012.1687. Epub 2012 Nov 6. Int J Oncol. 2013. PMID: 23135628 Free PMC article.
Cited by
-
Current relevance of hypoxia in head and neck cancer.Oncotarget. 2016 Aug 2;7(31):50781-50804. doi: 10.18632/oncotarget.9549. Oncotarget. 2016. PMID: 27434126 Free PMC article. Review.
-
ZBTB2 increases PDK4 expression by transcriptional repression of RelA/p65.Nucleic Acids Res. 2015 Feb 18;43(3):1609-25. doi: 10.1093/nar/gkv026. Epub 2015 Jan 21. Nucleic Acids Res. 2015. PMID: 25609694 Free PMC article.
-
Differing roles of pyruvate dehydrogenase kinases during mouse oocyte maturation.J Cell Sci. 2015 Jul 1;128(13):2319-29. doi: 10.1242/jcs.167049. Epub 2015 May 19. J Cell Sci. 2015. PMID: 25991547 Free PMC article.
-
Targeting pancreatic cancer with combinatorial treatment of CPI-613 and inhibitors of lactate metabolism.PLoS One. 2022 Apr 22;17(4):e0266601. doi: 10.1371/journal.pone.0266601. eCollection 2022. PLoS One. 2022. PMID: 35452495 Free PMC article.
-
Deciphering squamous cell carcinoma using multidimensional genomic approaches.J Skin Cancer. 2011;2011:541405. doi: 10.1155/2011/541405. Epub 2010 Dec 27. J Skin Cancer. 2011. PMID: 21234096 Free PMC article.
References
-
- Warburg, O. (1930) The Metabolism of Tumors, Constable Press, London, UK
-
- Deberardinis, R. J., Lum, J. J., Hatzivassiliou, G., and Thompson, C. B. (2008) Cell Metab. 7 11-20 - PubMed
-
- Harris, A. L. (2002) Nat. Rev. Cancer 2 38-47 - PubMed
-
- Mabjeesh, N. J., and Amir, S. (2007) Histol. Histopathol. 22 559-572 - PubMed
-
- Walenta, S., Schroeder, T., and Mueller-Klieser, W. (2004) Curr. Med. Chem. 11 2195-2204 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
