

Synaptotagmin VII regulates bone remodeling by modulating osteoclast and osteoblast secretion
- PMID: 18539119
- PMCID: PMC2480494
- DOI: 10.1016/j.devcel.2008.03.022
Synaptotagmin VII regulates bone remodeling by modulating osteoclast and osteoblast secretion
Abstract
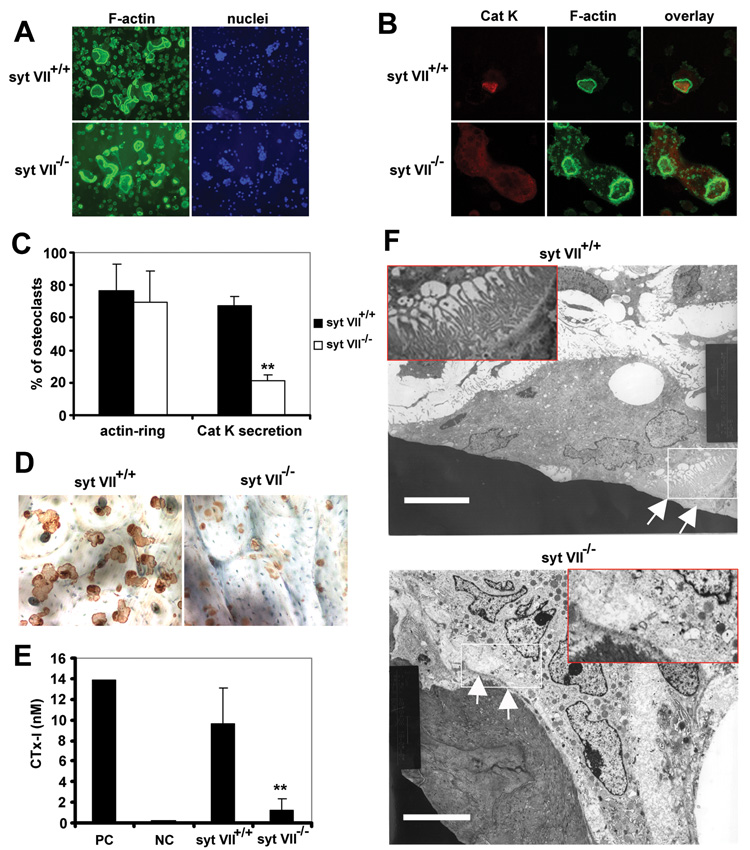
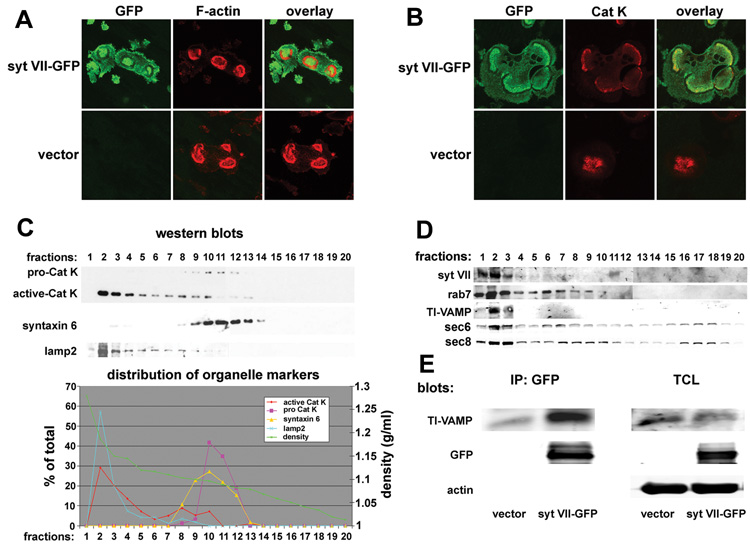
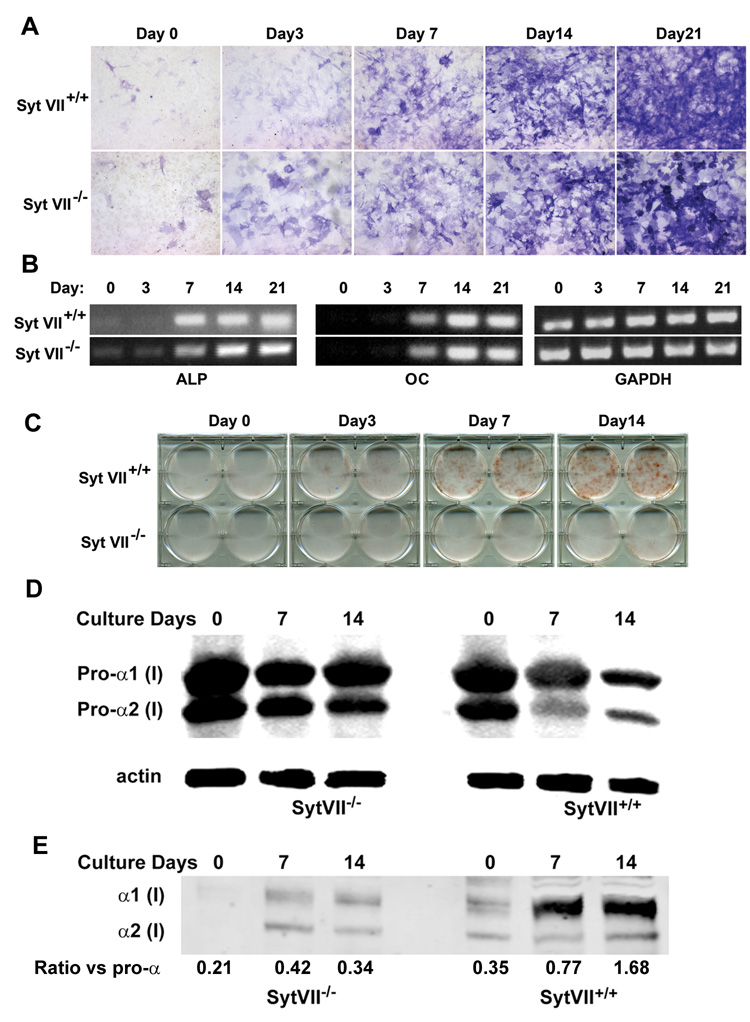
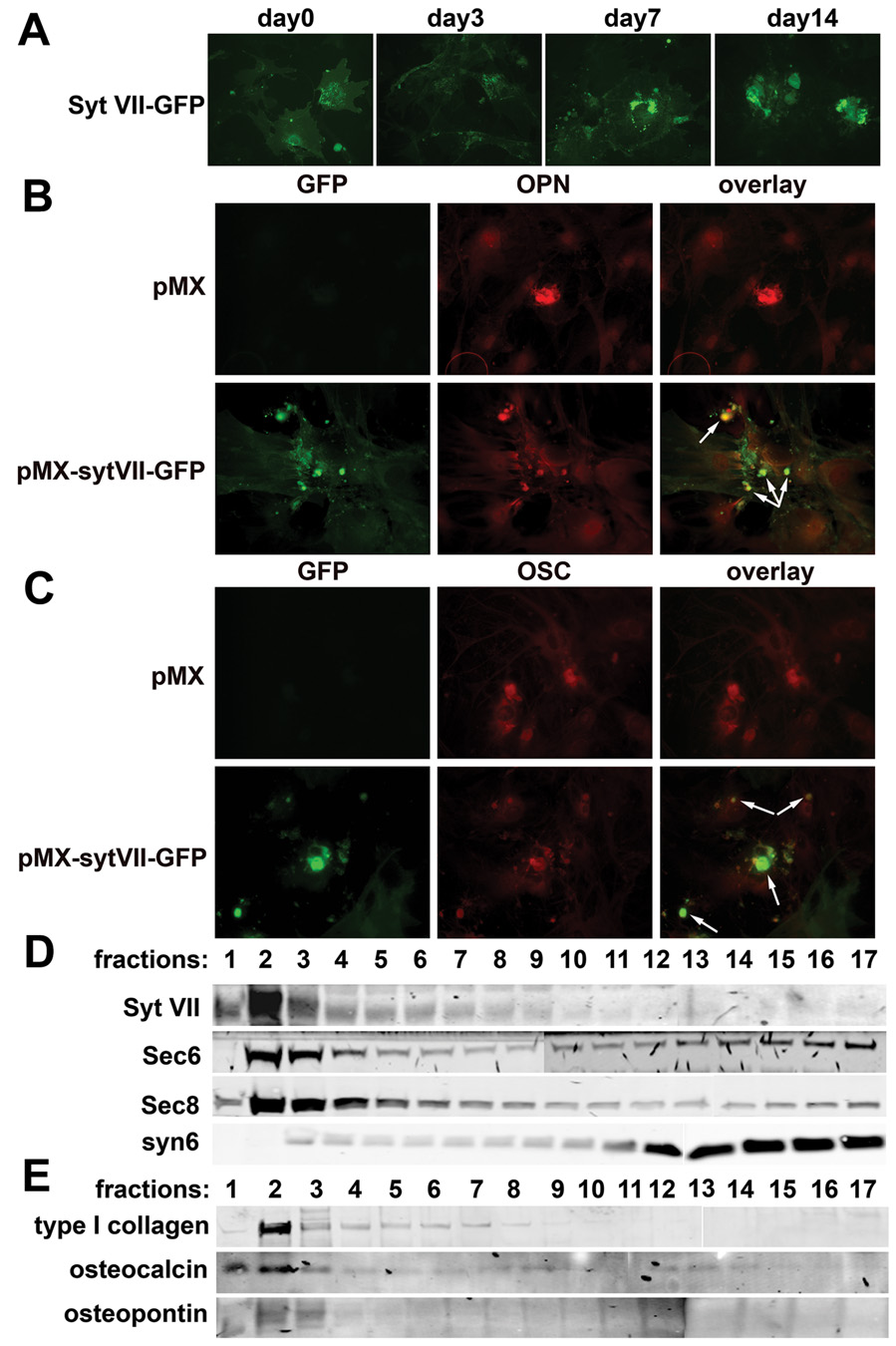
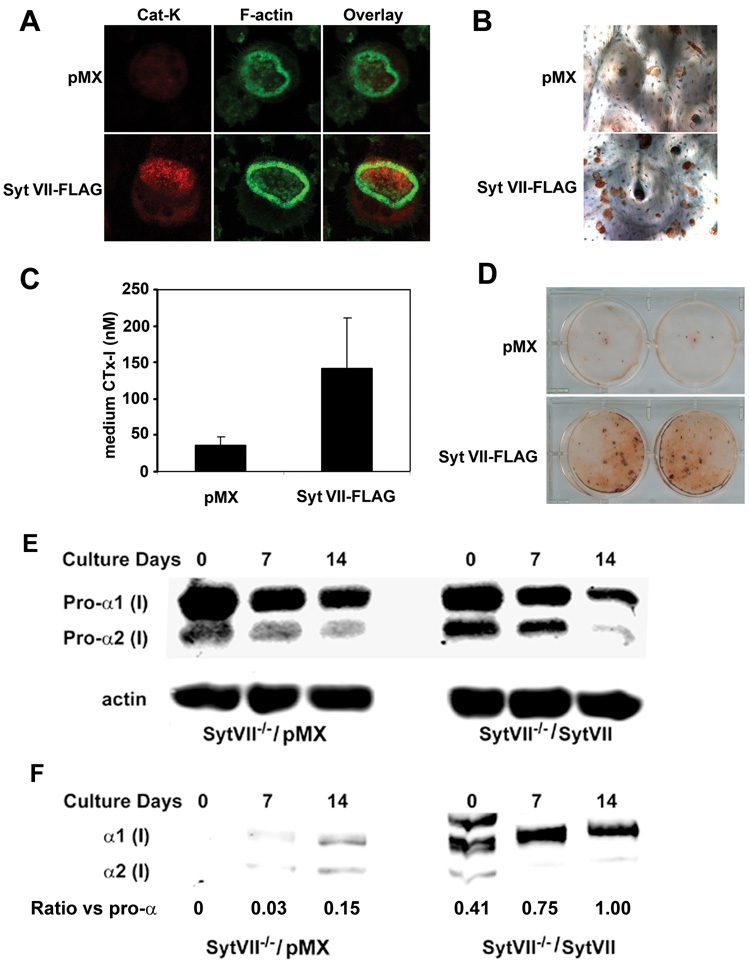
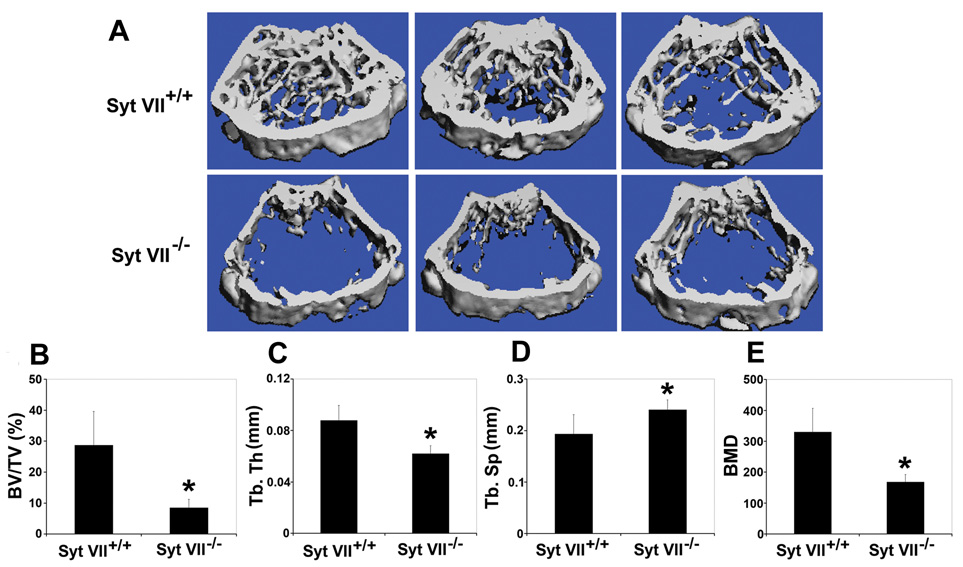
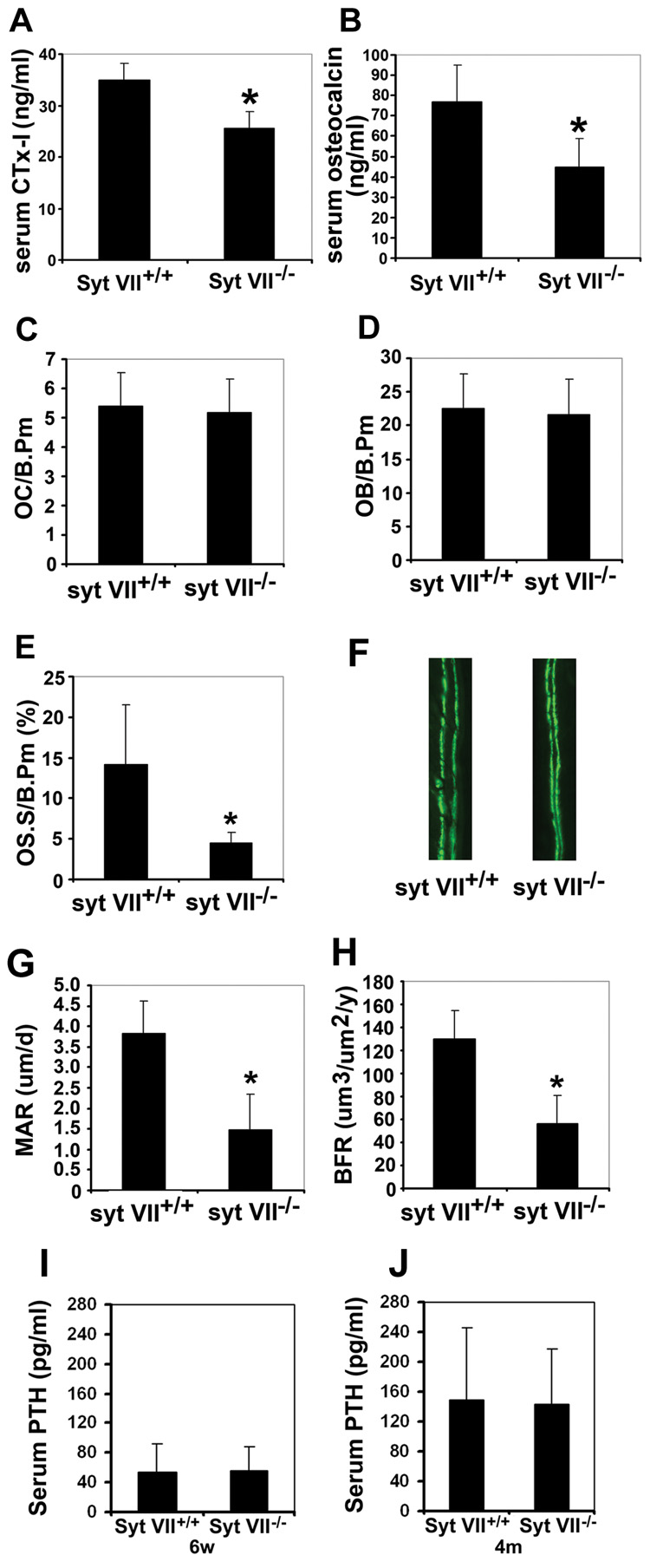
Maintenance of bone mass and integrity requires a tight balance between resorption by osteoclasts and formation by osteoblasts. Exocytosis of functional proteins is a prerequisite for the activity of both cells. In the present study, we show that synaptotagmin VII, a calcium sensor protein that regulates exocytosis, is associated with lysosomes in osteoclasts and bone matrix protein-containing vesicles in osteoblasts. Absence of synaptotagmin VII inhibits cathepsin K secretion and formation of the ruffled border in osteoclasts and bone matrix protein deposition in osteoblasts, without affecting the differentiation of either cell. Reflecting these in vitro findings, synaptotagmin VII-deficient mice are osteopenic due to impaired bone resorption and formation. Therefore, synaptotagmin VII plays an important role in bone remodeling and homeostasis by modulating secretory pathways functionally important in osteoclasts and osteoblasts.
Figures
Similar articles
-
Protein kinase C-delta deficiency perturbs bone homeostasis by selective uncoupling of cathepsin K secretion and ruffled border formation in osteoclasts.J Bone Miner Res. 2012 Dec;27(12):2452-63. doi: 10.1002/jbmr.1701. J Bone Miner Res. 2012. PMID: 22806935 Free PMC article.
-
Biglycan deficiency increases osteoclast differentiation and activity due to defective osteoblasts.Bone. 2006 Jun;38(6):778-86. doi: 10.1016/j.bone.2005.11.005. Epub 2005 Dec 20. Bone. 2006. PMID: 16364709
-
How do bone cells secrete proteins?Adv Exp Med Biol. 2010;658:105-9. doi: 10.1007/978-1-4419-1050-9_11. Adv Exp Med Biol. 2010. PMID: 19950020
-
There's more to life than neurotransmission: the regulation of exocytosis by synaptotagmin VII.Trends Cell Biol. 2005 Nov;15(11):626-31. doi: 10.1016/j.tcb.2005.09.001. Epub 2005 Sep 15. Trends Cell Biol. 2005. PMID: 16168654 Review.
-
Extracellular matrix networks in bone remodeling.Int J Biochem Cell Biol. 2015 Aug;65:20-31. doi: 10.1016/j.biocel.2015.05.008. Epub 2015 May 18. Int J Biochem Cell Biol. 2015. PMID: 25997875 Review.
Cited by
-
Protein kinase C-delta deficiency perturbs bone homeostasis by selective uncoupling of cathepsin K secretion and ruffled border formation in osteoclasts.J Bone Miner Res. 2012 Dec;27(12):2452-63. doi: 10.1002/jbmr.1701. J Bone Miner Res. 2012. PMID: 22806935 Free PMC article.
-
Sugar transporter Slc37a2 regulates bone metabolism in mice via a tubular lysosomal network in osteoclasts.Nat Commun. 2023 Feb 21;14(1):906. doi: 10.1038/s41467-023-36484-2. Nat Commun. 2023. PMID: 36810735 Free PMC article.
-
RUFY4 deletion prevents pathological bone loss by blocking endo-lysosomal trafficking of osteoclasts.Bone Res. 2024 May 15;12(1):29. doi: 10.1038/s41413-024-00326-8. Bone Res. 2024. PMID: 38744829 Free PMC article.
-
Non-Vesicular Release of Alarmin Prothymosin α Complex Associated with Annexin-2 Flop-Out.Cells. 2023 Jun 6;12(12):1569. doi: 10.3390/cells12121569. Cells. 2023. PMID: 37371039 Free PMC article. Review.
-
A non-immunological role for γ-interferon-inducible lysosomal thiol reductase (GILT) in osteoclastic bone resorption.Sci Adv. 2021 Apr 23;7(17):eabd3684. doi: 10.1126/sciadv.abd3684. Print 2021 Apr. Sci Adv. 2021. PMID: 33893096 Free PMC article.
References
-
- Anderson HC. Molecular biology of matrix vesicles. Clin Orthop. 1995;314:266–280. - PubMed
-
- Anderson HC, Garimella R, Tague SE. The role of matrix vesicles in growth plate development and biomineralization. Front Biosci. 2005;10:822–837. - PubMed
-
- Andrews NW, Chakrabarti S. There's more to life than neurotransmission: the regulation of exocytosis by synaptotagmin VII. Trends Cell Biol. 2005;15:626–631. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AR032788/AR/NIAMS NIH HHS/United States
- R01 AR046523/AR/NIAMS NIH HHS/United States
- R37 AR046523/AR/NIAMS NIH HHS/United States
- AR046523/AR/NIAMS NIH HHS/United States
- R01 AR046852/AR/NIAMS NIH HHS/United States
- R01 AR032788/AR/NIAMS NIH HHS/United States
- R01 AR046523-10/AR/NIAMS NIH HHS/United States
- R01 GM064625/GM/NIGMS NIH HHS/United States
- R01 AR032788-23/AR/NIAMS NIH HHS/United States
- R01 AR046852-09/AR/NIAMS NIH HHS/United States
- R21 AR054190-02/AR/NIAMS NIH HHS/United States
- AR046852/AR/NIAMS NIH HHS/United States
- AR054190/AR/NIAMS NIH HHS/United States
- R21 AR054190/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
