

Mitochondrial defects in acute multiple sclerosis lesions
- PMID: 18515320
- PMCID: PMC2442422
- DOI: 10.1093/brain/awn105
Mitochondrial defects in acute multiple sclerosis lesions
Abstract
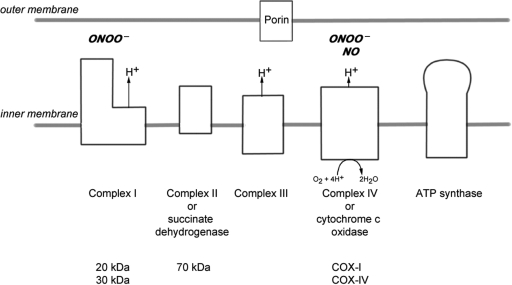
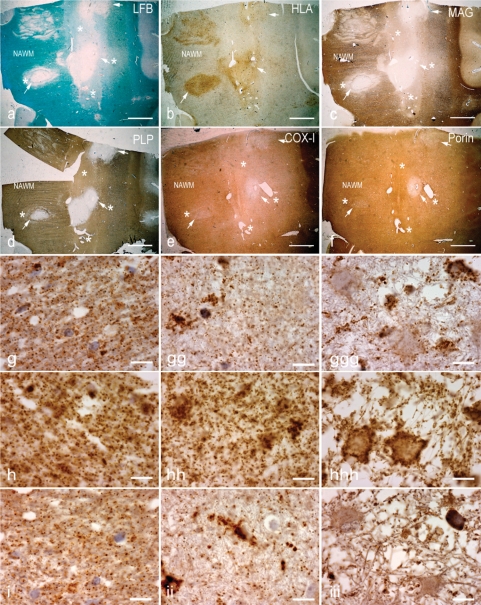
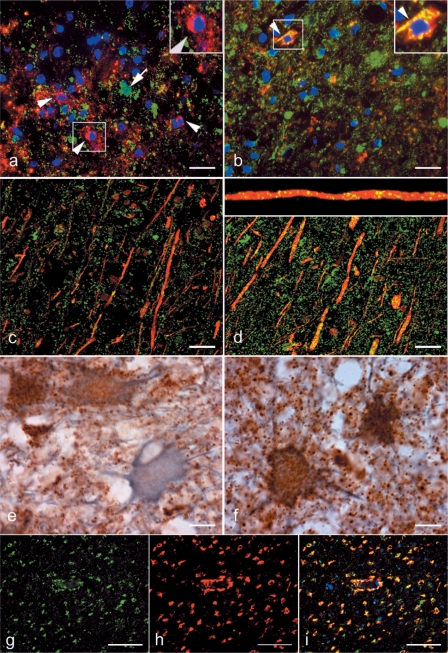
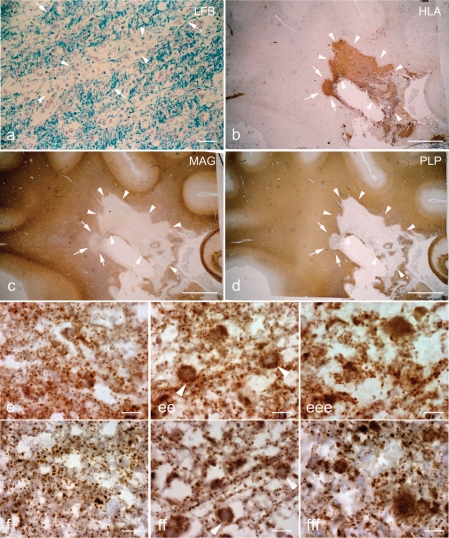
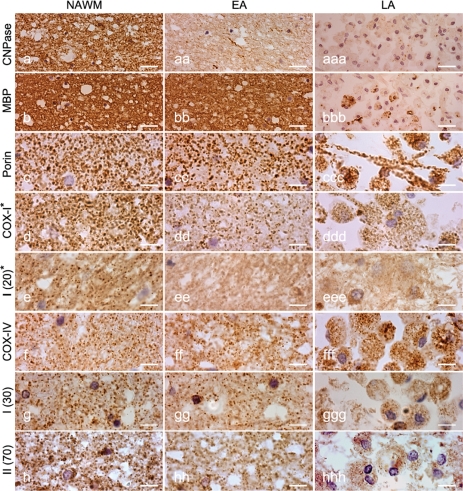
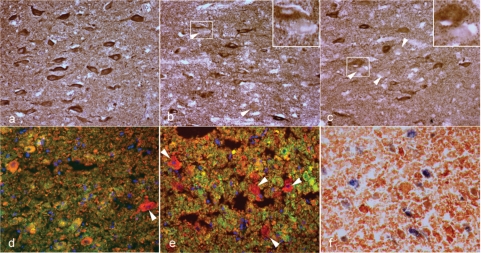
Multiple sclerosis is a chronic inflammatory disease, which leads to focal plaques of demyelination and tissue injury in the CNS. The structural and immunopathological patterns of demyelination suggest that different immune mechanisms may be involved in tissue damage. In a subtype of lesions, which are mainly found in patients with acute fulminant multiple sclerosis with Balo's type concentric sclerosis and in a subset of early relapsing remitting multiple sclerosis, the initial myelin changes closely resemble those seen in white matter stroke (WMS), suggesting a hypoxia-like tissue injury. Since mitochondrial injury may be involved in the pathogenesis of such lesions, we analysed a number of mitochondrial respiratory chain proteins in active lesions from acute multiple sclerosis and from WMS using immunohistochemistry. Functionally important defects of mitochondrial respiratory chain complex IV [cytochrome c oxidase (COX)] including its catalytic component (COX-I) are present in Pattern III but not in Pattern II multiple sclerosis lesions. The lack of immunohistochemically detected COX-I is apparent in oligodendrocytes, hypertrophied astrocytes and axons, but not in microglia. The profile of immunohistochemically detected mitochondrial respiratory chain complex subunits differs between multiple sclerosis and WMS. The findings suggest that hypoxia-like tissue injury in Pattern III multiple sclerosis lesions may be due to mitochondrial impairment.
Figures
Similar articles
-
NADPH oxidase expression in active multiple sclerosis lesions in relation to oxidative tissue damage and mitochondrial injury.Brain. 2012 Mar;135(Pt 3):886-99. doi: 10.1093/brain/aws012. Brain. 2012. PMID: 22366799 Free PMC article.
-
Tissue preconditioning may explain concentric lesions in Baló's type of multiple sclerosis.Brain. 2005 May;128(Pt 5):979-87. doi: 10.1093/brain/awh457. Epub 2005 Mar 17. Brain. 2005. PMID: 15774507
-
Mitochondrial changes within axons in multiple sclerosis.Brain. 2009 May;132(Pt 5):1161-74. doi: 10.1093/brain/awp046. Epub 2009 Mar 17. Brain. 2009. PMID: 19293237 Free PMC article.
-
[Recent progress in multiple sclerosis research: astrocytopathy in demyelinating diseases].Rinsho Shinkeigaku. 2010 Nov;50(11):788-93. doi: 10.5692/clinicalneurol.50.788. Rinsho Shinkeigaku. 2010. PMID: 21921443 Review. Japanese.
-
Astrocytopathy in Balo's disease.Mult Scler. 2011 Jul;17(7):771-9. doi: 10.1177/1352458511400475. Epub 2011 Apr 1. Mult Scler. 2011. PMID: 21459811 Review.
Cited by
-
Proteomic analysis of cerebrospinal fluid in a fulminant case of multiple sclerosis.Int J Mol Sci. 2012;13(6):7676-7693. doi: 10.3390/ijms13067676. Epub 2012 Jun 21. Int J Mol Sci. 2012. PMID: 22837721 Free PMC article.
-
NADPH oxidase expression in active multiple sclerosis lesions in relation to oxidative tissue damage and mitochondrial injury.Brain. 2012 Mar;135(Pt 3):886-99. doi: 10.1093/brain/aws012. Brain. 2012. PMID: 22366799 Free PMC article.
-
Food, nutrigenomics, and neurodegeneration--neuroprotection by what you eat!Mol Neurobiol. 2013 Oct;48(2):353-62. doi: 10.1007/s12035-013-8498-3. Epub 2013 Jun 28. Mol Neurobiol. 2013. PMID: 23813102 Review.
-
Sodium MRI in Multiple Sclerosis is Compatible with Intracellular Sodium Accumulation and Inflammation-Induced Hyper-Cellularity of Acute Brain Lesions.Sci Rep. 2016 Aug 10;6:31269. doi: 10.1038/srep31269. Sci Rep. 2016. PMID: 27507776 Free PMC article.
-
Mechanisms of neuronal dysfunction and degeneration in multiple sclerosis.Prog Neurobiol. 2011 Jan;93(1):1-12. doi: 10.1016/j.pneurobio.2010.09.005. Epub 2010 Oct 12. Prog Neurobiol. 2011. PMID: 20946934 Free PMC article. Review.
References
-
- Aboul-Enein F, Rauschka H, Kornek B, Stadelmann C, Stefferl A, Bruck W, et al. Preferential loss of myelin-associated glycoprotein reflects hypoxia-like white matter damage in stroke and inflammatory brain diseases. J Neuropathol Exp Neurol. 2003;62:25–33. - PubMed
-
- Almeida A, Moncada S, Bolanos JP. Nitric oxide switches on glycolysis through the AMP protein kinase and 6-phosphofructo-2-kinase pathway. Nat Cell Biol. 2004;6:45–51. - PubMed
-
- Barnett MH, Prineas JW. Relapsing and remitting multiple sclerosis: pathology of the newly forming lesion. Ann Neurol. 2004;55:458–68. - PubMed
-
- Bolanos JP, Almeida A, Stewart V, Peuchen S, Land JM, Clark JB, et al. Nitric oxide-mediated mitochondrial damage in the brain: mechanisms and implications for neurodegenerative diseases. J Neurochem. 1997;68:2227–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
