

Pharmacoproteomics of a metalloproteinase hydroxamate inhibitor in breast cancer cells: dynamics of membrane type 1 matrix metalloproteinase-mediated membrane protein shedding
- PMID: 18505826
- PMCID: PMC2493375
- DOI: 10.1128/MCB.01775-07
Pharmacoproteomics of a metalloproteinase hydroxamate inhibitor in breast cancer cells: dynamics of membrane type 1 matrix metalloproteinase-mediated membrane protein shedding
Abstract
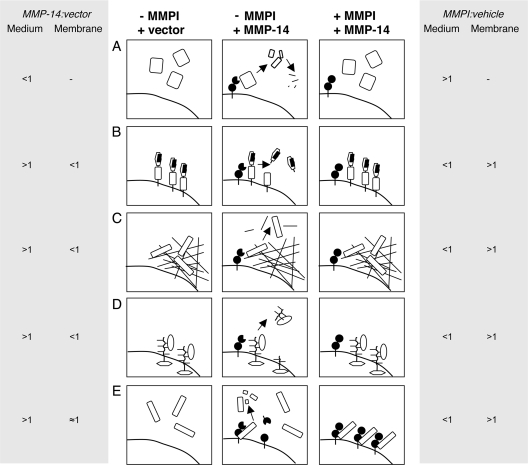
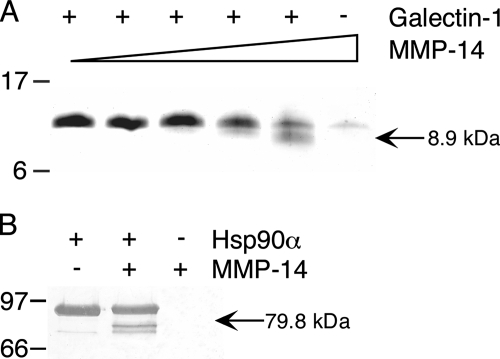
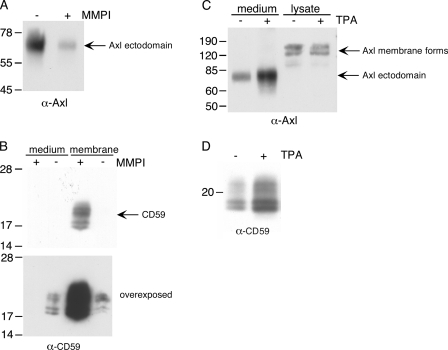
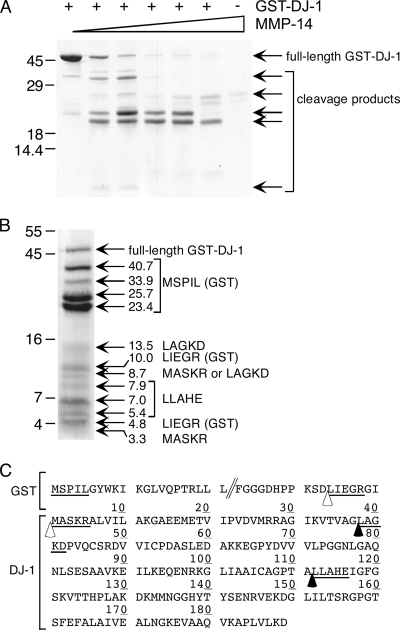
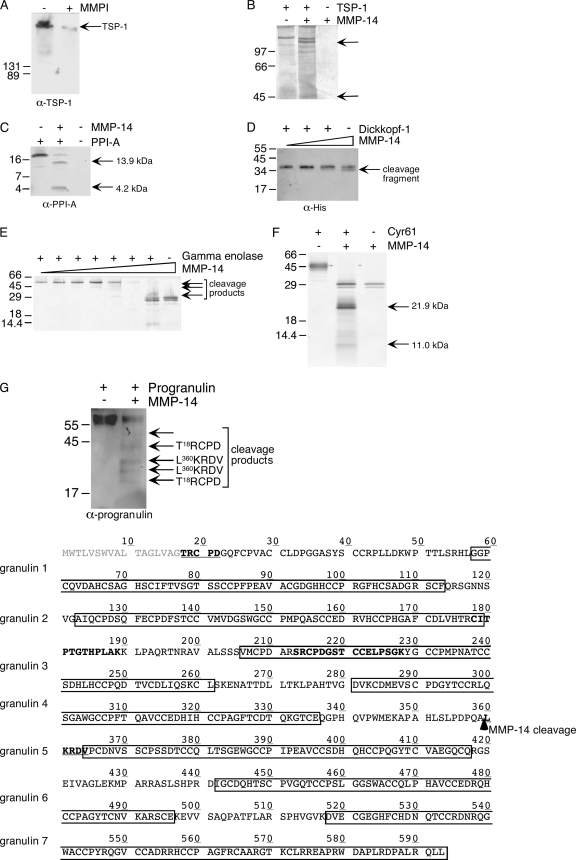
Broad-spectrum matrix metalloproteinase (MMP) inhibitors (MMPI) were unsuccessful in cancer clinical trials, partly due to side effects resulting from limited knowledge of the full repertoire of MMP substrates, termed the substrate degradome, and hence the in vivo functions of MMPs. To gain further insight into the degradome of MMP-14 (membrane type 1 MMP) an MMPI, prinomastat (drug code AG3340), was used to reduce proteolytic processing and ectodomain shedding in human MDA-MB-231 breast cancer cells transfected with MMP-14. We report a quantitative proteomic evaluation of the targets and effects of the inhibitor in this cell-based system. Proteins in cell-conditioned medium (the secretome) and membrane fractions with levels that were modulated by the MMPI were identified by isotope-coded affinity tag (ICAT) labeling and tandem mass spectrometry. Comparisons of the expression of MMP-14 with that of a vector control resulted in increased MMP-14/vector ICAT ratios for many proteins in conditioned medium, indicating MMP-14-mediated ectodomain shedding. Following MMPI treatment, the MMPI/vehicle ICAT ratio was reversed, suggesting that MMP-14-mediated shedding of these proteins was blocked by the inhibitor. The reduction in shedding or the release of substrates from pericellular sites in the presence of the MMPI was frequently accompanied by the accumulation of the protein in the plasma membrane, as indicated by high MMPI/vehicle ICAT ratios. Considered together, this is a strong predictor of biologically relevant substrates cleaved in the cellular context that led to the identification of many undescribed MMP-14 substrates, 20 of which we validated biochemically, including DJ-1, galectin-1, Hsp90alpha, pentraxin 3, progranulin, Cyr61, peptidyl-prolyl cis-trans isomerase A, and dickkopf-1. Other proteins with altered levels, such as Kunitz-type protease inhibitor 1 and beta-2-microglobulin, were not substrates in biochemical assays, suggesting an indirect affect of the MMPI, which might be important in drug development as biomarkers or, in preclinical phases, to predict systemic drug actions and adverse side effects. Hence, this approach describes the dynamic pattern of cell membrane ectodomain shedding and its perturbation upon metalloproteinase drug treatment.
Figures

Similar articles
-
Proteomic validation of protease drug targets: pharmacoproteomics of matrix metalloproteinase inhibitor drugs using isotope-coded affinity tag labelling and tandem mass spectrometry.Curr Pharm Des. 2007;13(3):263-70. doi: 10.2174/138161207779313524. Curr Pharm Des. 2007. PMID: 17313360 Review.
-
Proteomics discovery of metalloproteinase substrates in the cellular context by iTRAQ labeling reveals a diverse MMP-2 substrate degradome.Mol Cell Proteomics. 2007 Apr;6(4):611-23. doi: 10.1074/mcp.M600341-MCP200. Epub 2007 Jan 1. Mol Cell Proteomics. 2007. PMID: 17200105
-
Protease degradomics: mass spectrometry discovery of protease substrates and the CLIP-CHIP, a dedicated DNA microarray of all human proteases and inhibitors.Biol Chem. 2004 Jun;385(6):493-504. doi: 10.1515/BC.2004.058. Biol Chem. 2004. PMID: 15255181
-
Membrane protease proteomics: Isotope-coded affinity tag MS identification of undescribed MT1-matrix metalloproteinase substrates.Proc Natl Acad Sci U S A. 2004 May 4;101(18):6917-22. doi: 10.1073/pnas.0305862101. Epub 2004 Apr 26. Proc Natl Acad Sci U S A. 2004. PMID: 15118097 Free PMC article.
-
Degradomics: systems biology of the protease web. Pleiotropic roles of MMPs in cancer.Cancer Metastasis Rev. 2006 Mar;25(1):69-75. doi: 10.1007/s10555-006-7890-0. Cancer Metastasis Rev. 2006. PMID: 16680573 Review.
Cited by
-
Caught between a "Rho" and a hard place: are CCN1/CYR61 and CCN2/CTGF the arbiters of microvascular stiffness?J Cell Commun Signal. 2020 Mar;14(1):21-29. doi: 10.1007/s12079-019-00529-3. Epub 2019 Aug 2. J Cell Commun Signal. 2020. PMID: 31376071 Free PMC article. Review.
-
Progranulin: a novel regulator of gastrointestinal cancer progression.Transl Gastrointest Cancer. 2013 Jul;2(3):145-151. doi: 10.3978/j.issn.2224-4778.2013.02.02. Transl Gastrointest Cancer. 2013. PMID: 24040621 Free PMC article.
-
The lysosomal function of progranulin, a guardian against neurodegeneration.Acta Neuropathol. 2018 Jul;136(1):1-17. doi: 10.1007/s00401-018-1861-8. Epub 2018 May 9. Acta Neuropathol. 2018. PMID: 29744576 Free PMC article. Review.
-
Novel Insights into PARK7 (DJ-1), a Potential Anti-Cancer Therapeutic Target, and Implications for Cancer Progression.J Clin Med. 2020 Apr 26;9(5):1256. doi: 10.3390/jcm9051256. J Clin Med. 2020. PMID: 32357493 Free PMC article. Review.
-
The promotion of bone healing by progranulin, a downstream molecule of BMP-2, through interacting with TNF/TNFR signaling.Biomaterials. 2013 Sep;34(27):6412-21. doi: 10.1016/j.biomaterials.2013.05.030. Epub 2013 Jun 5. Biomaterials. 2013. PMID: 23746860 Free PMC article.
References
-
- Abdul, M., and N. Hoosein. 2000. Changes in beta-2 microglobulin expression in prostate cancer. Urol. Oncol. 5168-172. - PubMed
-
- Allain, F., C. Vanpouille, M. Carpentier, M. C. Slomianny, S. Durieux, and G. Spik. 2002. Interaction with glycosaminoglycans is required for cyclophilin B to trigger integrin-mediated adhesion of peripheral blood T lymphocytes to extracellular matrix. Proc. Natl. Acad. Sci. USA 992714-2719. - PMC - PubMed
-
- Allegrini, G., F. A. Goulette, J. W. Darnowski, and P. Calabresi. 2004. Thrombospondin-1 plus irinotecan: a novel antiangiogenic-chemotherapeutic combination that inhibits the growth of advanced human colon tumor xenografts in mice. Cancer Chemother. Pharmacol. 53261-266. - PubMed
-
- Andolfo, A., W. R. English, M. Resnati, G. Murphy, F. Blasi, and N. Sidenius. 2002. Metalloproteases cleave the urokinase-type plasminogen activator receptor in the D1-D2 linker region and expose epitopes not present in the intact soluble receptor. Thromb. Haemostasis 88298-306. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous