

Characterization of the major fragance gene from an aromatic japonica rice and analysis of its diversity in Asian cultivated rice
- PMID: 18491070
- PMCID: PMC2470208
- DOI: 10.1007/s00122-008-0780-9
Characterization of the major fragance gene from an aromatic japonica rice and analysis of its diversity in Asian cultivated rice
Abstract
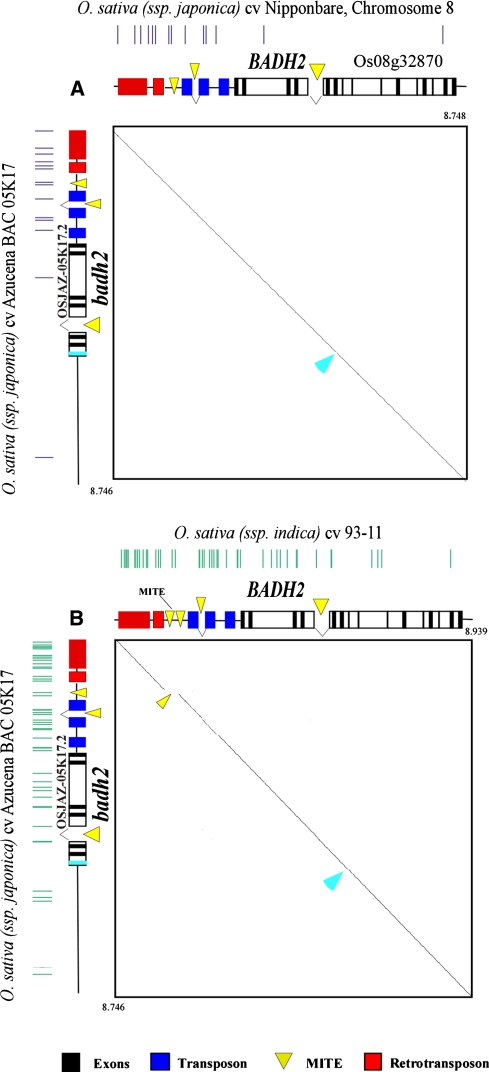
In Asian cultivated rice (Oryza sativa L.), aroma is one of the most valuable traits in grain quality and 2-ACP is the main volatile compound contributing to the characteristic popcorn-like odour of aromatic rices. Although the major locus for grain fragrance (frg gene) has been described recently in Basmati rice, this gene has not been characterised in true japonica varieties and molecular information available on the genetic diversity and evolutionary origin of this gene among the different varieties is still limited. Here we report on characterisation of the frg gene in the Azucena variety, one of the few aromatic japonica cultivars. We used a RIL population from a cross between Azucena and IR64, a non-aromatic indica, the reference genomic sequence of Nipponbare (japonica) and 93-11 (indica) as well as an Azucena BAC library, to identify the major fragance gene in Azucena. We thus identified a betaine aldehyde dehydrogenase gene, badh2, as the candidate locus responsible for aroma, which presented exactly the same mutation as that identified in Basmati and Jasmine-like rices. Comparative genomic analyses showed very high sequence conservation between Azucena and Nipponbare BADH2, and a MITE was identified in the promotor region of the BADH2 allele in 93-11. The badh2 mutation and MITE were surveyed in a representative rice collection, including traditional aromatic and non-aromatic rice varieties, and strongly suggested a monophylogenetic origin of this badh2 mutation in Asian cultivated rices. Altogether these new data are discussed here in the light of current hypotheses on the origin of rice genetic diversity.
Figures


Similar articles
-
The origin and evolution of fragrance in rice (Oryza sativa L.).Proc Natl Acad Sci U S A. 2009 Aug 25;106(34):14444-9. doi: 10.1073/pnas.0904077106. Epub 2009 Aug 17. Proc Natl Acad Sci U S A. 2009. PMID: 19706531 Free PMC article.
-
Whole genome sequencing of Oryza sativa L. cv. Seeragasamba identifies a new fragrance allele in rice.PLoS One. 2017 Nov 30;12(11):e0188920. doi: 10.1371/journal.pone.0188920. eCollection 2017. PLoS One. 2017. PMID: 29190814 Free PMC article.
-
Evaluation of Whole-Genome Sequence, Genetic Diversity, and Agronomic Traits of Basmati Rice (Oryza sativa L.).Front Genet. 2020 Feb 21;11:86. doi: 10.3389/fgene.2020.00086. eCollection 2020. Front Genet. 2020. PMID: 32153645 Free PMC article.
-
Thirty-three years of 2-acetyl-1-pyrroline, a principal basmati aroma compound in scented rice (Oryza sativa L.): a status review.J Sci Food Agric. 2017 Jan;97(2):384-395. doi: 10.1002/jsfa.7875. Epub 2016 Jul 26. J Sci Food Agric. 2017. PMID: 27376959 Review.
-
CRISPR-Cas9-mediated editing of BADH2 gene triggered fragrance revolution in rice.Physiol Plant. 2023 Jan;175(1):e13871. doi: 10.1111/ppl.13871. Physiol Plant. 2023. PMID: 36748269 Review.
Cited by
-
Proteomic Analysis of Isogenic Rice Reveals Proteins Correlated with Aroma Compound Biosynthesis at Different Developmental Stages.Mol Biotechnol. 2016 Feb;58(2):117-29. doi: 10.1007/s12033-015-9906-x. Mol Biotechnol. 2016. PMID: 26646386
-
A PCR-based marker for a locus conferring the aroma in Myanmar rice (Oryza sativa L.).Theor Appl Genet. 2012 Sep;125(5):887-96. doi: 10.1007/s00122-012-1880-0. Epub 2012 May 11. Theor Appl Genet. 2012. PMID: 22576235
-
Resequencing Reveals Different Domestication Rate for BADH1 and BADH2 in Rice (Oryza sativa).PLoS One. 2015 Aug 10;10(8):e0134801. doi: 10.1371/journal.pone.0134801. eCollection 2015. PLoS One. 2015. PMID: 26258482 Free PMC article.
-
Effects of paclobutrazol seed priming on seedling quality, photosynthesis, and physiological characteristics of fragrant rice.BMC Plant Biol. 2024 Jan 17;24(1):53. doi: 10.1186/s12870-023-04683-0. BMC Plant Biol. 2024. PMID: 38229011 Free PMC article.
-
Prospects of breeding high-quality rice using post-genomic tools.Theor Appl Genet. 2015 Aug;128(8):1449-66. doi: 10.1007/s00122-015-2537-6. Epub 2015 May 21. Theor Appl Genet. 2015. PMID: 25993897 Review.
References
-
- Amarawathi Y, Singh R, Sing AK, Singh VP, Mohapatra T, Sharma TR, Singh NK (2007) Mapping ofd quantitative trait loci for basmati quality traits in rice (Oryza sativa L.). Mol Breed. doi:10.1007/s11032-007-9108-8
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
