

Synaptic vesicle protein trafficking at the glutamate synapse
- PMID: 18472224
- PMCID: PMC2667334
- DOI: 10.1016/j.neuroscience.2008.03.029
Synaptic vesicle protein trafficking at the glutamate synapse
Abstract
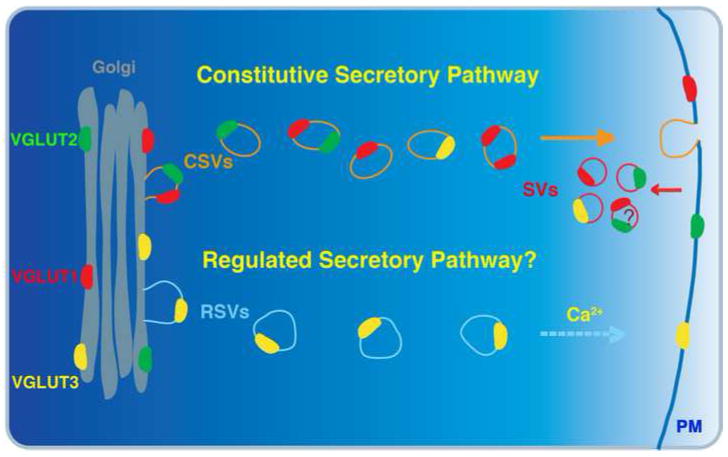
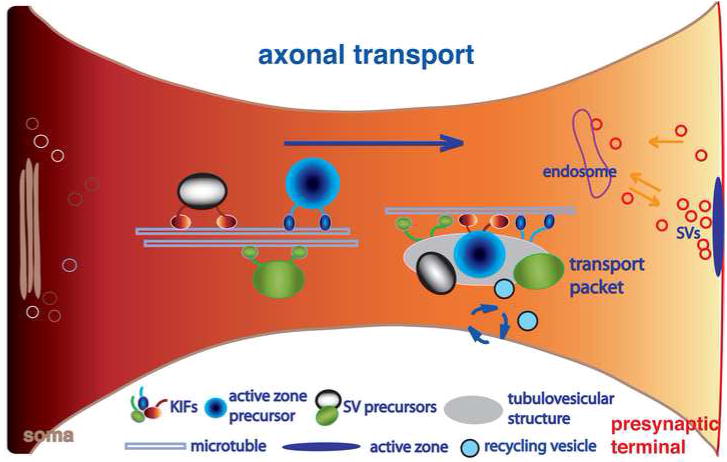
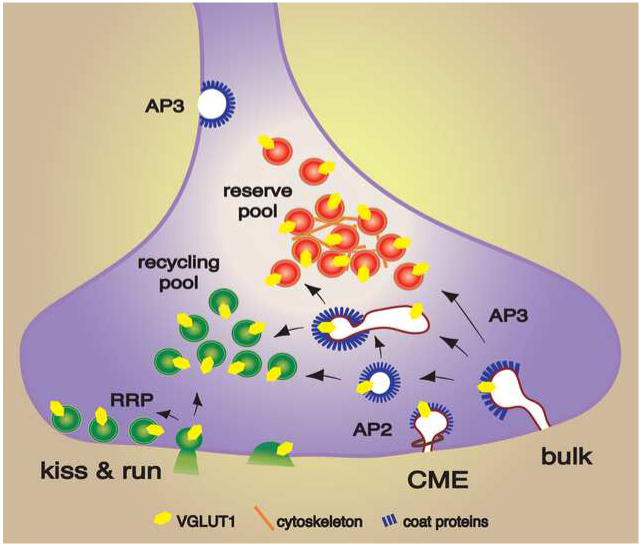
Expression of the integral and associated proteins of synaptic vesicles is subject to regulation over time, by region, and in response to activity. The process by which changes in protein levels and isoforms result in different properties of neurotransmitter release involves protein trafficking to the synaptic vesicle. How newly synthesized proteins are incorporated into synaptic vesicles at the presynaptic bouton is poorly understood. During synaptogenesis, synaptic vesicle proteins sort through the secretory pathway and are transported down the axon in precursor vesicles that undergo maturation to form synaptic vesicles. Changes in protein content of synaptic vesicles could involve the formation of new vesicles that either mix with the previous complement of vesicles or replace them, presumably by their degradation or inactivation. Alternatively, new proteins could individually incorporate into existing synaptic vesicles, changing their functional properties. Glutamatergic vesicles likely express many of the same integral membrane proteins and share certain common mechanisms of biogenesis, recycling, and degradation with other synaptic vesicles. However, glutamatergic vesicles are defined by their ability to package glutamate for release, a property conferred by the expression of a vesicular glutamate transporter (VGLUT). VGLUTs are subject to regional, developmental, and activity-dependent changes in expression. In addition, VGLUT isoforms differ in their trafficking, which may target them to different pathways during biogenesis or after recycling, which may in turn sort them to different vesicle pools. Emerging data indicate that differences in the association of VGLUTs and other synaptic vesicle proteins with endocytic adaptors may influence their trafficking. These observations indicate that independent regulation of synaptic vesicle protein trafficking has the potential to influence synaptic vesicle protein composition, the maintenance of synaptic vesicle pools, and the release of glutamate in response to changing physiological requirements.
Figures

Similar articles
-
VGLUTs: 'exciting' times for glutamatergic research?Neurosci Res. 2006 Aug;55(4):343-51. doi: 10.1016/j.neures.2006.04.016. Epub 2006 Jun 9. Neurosci Res. 2006. PMID: 16765470 Review.
-
Complementary distribution of vesicular glutamate transporters in the central nervous system.Neurosci Res. 2002 Apr;42(4):243-50. doi: 10.1016/s0168-0102(02)00009-3. Neurosci Res. 2002. PMID: 11985876 Review.
-
Rustling synaptic vesicle cargo after exocytosis.Neuron. 2006 Jul 6;51(1):3-5. doi: 10.1016/j.neuron.2006.06.015. Neuron. 2006. PMID: 16815324 Review.
-
Spike Activity Regulates Vesicle Filling at a Glutamatergic Synapse.J Neurosci. 2020 Jun 24;40(26):4972-4980. doi: 10.1523/JNEUROSCI.2945-19.2020. Epub 2020 May 19. J Neurosci. 2020. PMID: 32430294 Free PMC article.
-
The synaptic vesicle cycle.Annu Rev Neurosci. 2004;27:509-47. doi: 10.1146/annurev.neuro.26.041002.131412. Annu Rev Neurosci. 2004. PMID: 15217342 Review.
Cited by
-
Protein sorting, targeting and trafficking in photoreceptor cells.Prog Retin Eye Res. 2013 Sep;36:24-51. doi: 10.1016/j.preteyeres.2013.03.002. Epub 2013 Apr 3. Prog Retin Eye Res. 2013. PMID: 23562855 Free PMC article. Review.
-
Sorting of the vesicular GABA transporter to functional vesicle pools by an atypical dileucine-like motif.J Neurosci. 2013 Jun 26;33(26):10634-46. doi: 10.1523/JNEUROSCI.0329-13.2013. J Neurosci. 2013. PMID: 23804087 Free PMC article.
-
Analysis of synaptic gene expression in the neocortex of primates reveals evolutionary changes in glutamatergic neurotransmission.Cereb Cortex. 2015 Jun;25(6):1596-607. doi: 10.1093/cercor/bht354. Epub 2014 Jan 9. Cereb Cortex. 2015. PMID: 24408959 Free PMC article.
-
Pathways for Emotions: Specializations in the Amygdalar, Mediodorsal Thalamic, and Posterior Orbitofrontal Network.J Neurosci. 2015 Aug 26;35(34):11976-87. doi: 10.1523/JNEUROSCI.2157-15.2015. J Neurosci. 2015. PMID: 26311778 Free PMC article.
-
Postsynaptic VAMP/Synaptobrevin Facilitates Differential Vesicle Trafficking of GluA1 and GluA2 AMPA Receptor Subunits.PLoS One. 2015 Oct 21;10(10):e0140868. doi: 10.1371/journal.pone.0140868. eCollection 2015. PLoS One. 2015. PMID: 26488171 Free PMC article.
References
-
- Ahmari SE, Buchanan J, Smith SJ. Assembly of presynaptic active zones from cytoplasmic transport packets. Nat Neurosci. 2000;3:445–451. - PubMed
-
- Aihara Y, Mashima H, Onda H, Hisano S, Kasuya H, Hori T, Yamada S, Tomura H, Yamada Y, Inoue I, Kojima I, Takeda J. Molecular cloning of a novel brain-type Na(+)-dependent inorganic phosphate cotransporter. J Neurochem. 2000;74:2622–2625. - PubMed
-
- Altrock WD, tom Dieck S, Sokolov M, Meyer AC, Sigler A, Brakebusch C, Fassler R, Richter K, Boeckers TM, Potschka H, Brandt C, Loscher W, Grimberg D, Dresbach T, Hempelmann A, Hassan H, Balschun D, Frey JU, Brandstatter JH, Garner CC, Rosenmund C, Gundelfinger ED. Functional inactivation of a fraction of excitatory synapses in mice deficient for the active zone protein bassoon. Neuron. 2003;37:787–800. - PubMed
-
- Antonov I, Chang S, Zakharenko S, Popov SV. Distribution of neurotransmitter secretion in growing axons. Neuroscience. 1999;90:975–984. - PubMed
-
- Ashton AC, Ushkaryov YA. Properties of synaptic vesicle pools in mature central nerve terminals. J Biol Chem. 2005;280:37278–37288. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
