

The cyclic AMP effector Epac integrates pro- and anti-fibrotic signals
- PMID: 18434542
- PMCID: PMC2359804
- DOI: 10.1073/pnas.0801490105
The cyclic AMP effector Epac integrates pro- and anti-fibrotic signals
Erratum in
- Proc Natl Acad Sci U S A. 2008 Jun 10;105(23):8160
Abstract
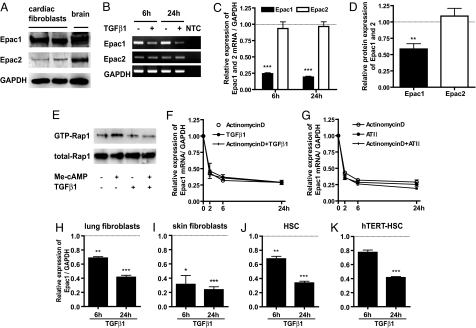
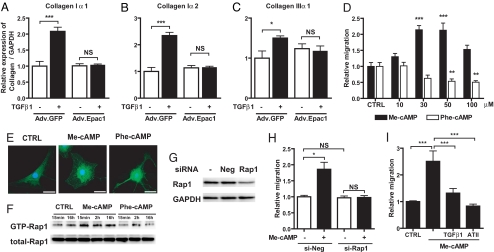
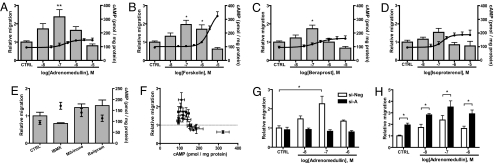
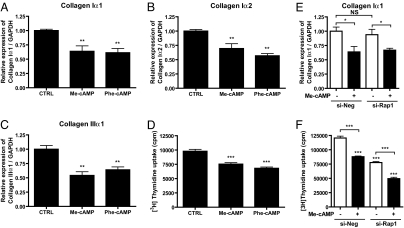
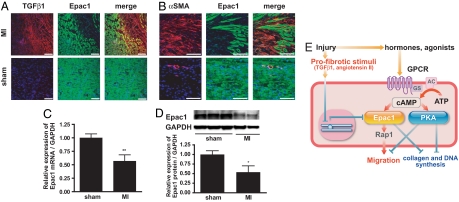
Scar formation occurs during the late stages of the inflammatory response but, when excessive, produces fibrosis that can lead to functional and structural damage of tissues. Here, we show that the profibrogenic agonist, transforming growth factor beta1, transcriptionally decreases expression of Exchange protein activated by cAMP 1 (Epac1) in fibroblasts/fibroblast-like cells from multiple tissues (i.e., cardiac, lung, and skin fibroblasts and hepatic stellate cells). Overexpression of Epac1 inhibits transforming growth factor beta1-induced collagen synthesis, indicating that a decrease of Epac1 expression appears to be necessary for the fibrogenic phenotype, an idea supported by evidence that Epac1 expression in cardiac fibroblasts is inhibited after myocardial infarction. Epac and protein kinase A, a second mediator of cAMP action, have opposite effects on migration but both inhibit synthesis of collagen and DNA by fibroblasts. Epac is preferentially activated by low concentrations of cAMP and stimulates migration via the small G protein Rap1 but inhibits collagen synthesis in a Rap1-independent manner. The regulation of Epac expression and activation thus appear to be critical for the integration of pro- and anti-fibrotic signals and for the regulation of fibroblast function.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
EPAC activation inhibits acetaldehyde-induced activation and proliferation of hepatic stellate cell via Rap1.Can J Physiol Pharmacol. 2016 May;94(5):498-507. doi: 10.1139/cjpp-2015-0437. Epub 2015 Nov 19. Can J Physiol Pharmacol. 2016. PMID: 26854595
-
Small GTPase Rap1 regulates cell migration through regulation of small GTPase RhoA activity in response to transforming growth factor-β1.J Cell Physiol. 2013 Nov;228(11):2119-26. doi: 10.1002/jcp.24383. J Cell Physiol. 2013. PMID: 23559363
-
Rap1-mediated activation of extracellular signal-regulated kinases by cyclic AMP is dependent on the mode of Rap1 activation.Mol Cell Biol. 2006 Mar;26(6):2130-45. doi: 10.1128/MCB.26.6.2130-2145.2006. Mol Cell Biol. 2006. PMID: 16507992 Free PMC article.
-
Role of the cAMP-binding protein Epac in cardiovascular physiology and pathophysiology.Pflugers Arch. 2010 Mar;459(4):535-46. doi: 10.1007/s00424-009-0747-y. Epub 2009 Oct 25. Pflugers Arch. 2010. PMID: 19855995 Review.
-
cAMP and Epac in the regulation of tissue fibrosis.Br J Pharmacol. 2012 May;166(2):447-56. doi: 10.1111/j.1476-5381.2012.01847.x. Br J Pharmacol. 2012. PMID: 22233238 Free PMC article. Review.
Cited by
-
Isoform-specific antagonists of exchange proteins directly activated by cAMP.Proc Natl Acad Sci U S A. 2012 Nov 6;109(45):18613-8. doi: 10.1073/pnas.1210209109. Epub 2012 Oct 22. Proc Natl Acad Sci U S A. 2012. PMID: 23091014 Free PMC article.
-
Cardiac cAMP-PKA Signaling Compartmentalization in Myocardial Infarction.Cells. 2021 Apr 16;10(4):922. doi: 10.3390/cells10040922. Cells. 2021. PMID: 33923648 Free PMC article. Review.
-
Cyclic GMP kinase and RhoA Ser188 phosphorylation integrate pro- and antifibrotic signals in blood vessels.Mol Cell Biol. 2009 Nov;29(22):6018-32. doi: 10.1128/MCB.00225-09. Epub 2009 Sep 8. Mol Cell Biol. 2009. PMID: 19737918 Free PMC article.
-
cAMP Signalling Pathway in Biocontrol Fungi.Curr Issues Mol Biol. 2022 Jun 4;44(6):2622-2634. doi: 10.3390/cimb44060179. Curr Issues Mol Biol. 2022. PMID: 35735620 Free PMC article. Review.
-
Increase in cellular cyclic AMP concentrations reverses the profibrogenic phenotype of cardiac myofibroblasts: a novel therapeutic approach for cardiac fibrosis.Mol Pharmacol. 2013 Dec;84(6):787-93. doi: 10.1124/mol.113.087742. Epub 2013 Oct 1. Mol Pharmacol. 2013. PMID: 24085841 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
