

Review
doi: 10.1038/nrmicro1890.
Modification of intracellular membrane structures for virus replication
Affiliations
- PMID: 18414501
- PMCID: PMC7096853
- DOI: 10.1038/nrmicro1890
Item in Clipboard
Review
Modification of intracellular membrane structures for virus replication
Nat Rev Microbiol.
2008 May.
Abstract
Viruses are intracellular parasites that use the host cell they infect to produce new infectious progeny. Distinct steps of the virus life cycle occur in association with the cytoskeleton or cytoplasmic membranes, which are often modified during infection. Plus-stranded RNA viruses induce membrane proliferations that support the replication of their genomes. Similarly, cytoplasmic replication of some DNA viruses occurs in association with modified cellular membranes. We describe how viruses modify intracellular membranes, highlight similarities between the structures that are induced by viruses of different families and discuss how these structures could be formed.
Figures
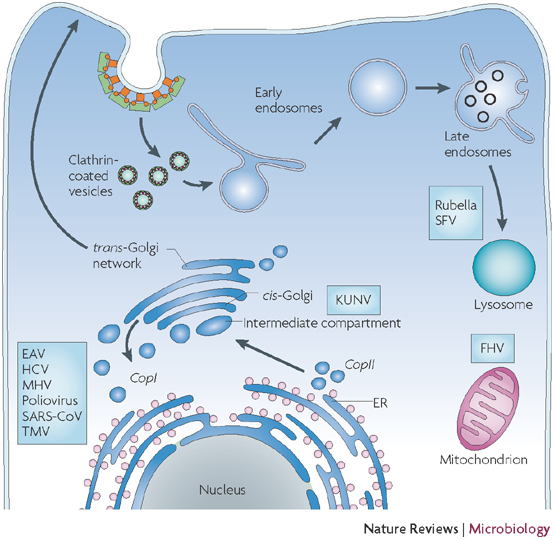
Schematic representation of a cell and different intracellular organelles. Proteins that are destined for secretion enter the secretory pathway by co-translational translocation into the endoplasmic reticulum (ER) (pink dots represent ribosomes). These proteins are then transported in a coatomer protein complex (COP) II-dependent way to the Golgi complex in a process that probably involves COPII-coated vesicles and membrane structures that are located in the intermediate compartment between the ER and the Golgi complex. Proteins can be recycled back to the ER using COPI-coated vesicles or can be transported through the Golgi complex. At the trans–Golgi network, they leave the Golgi and are transported to the plasma membrane. Endocytosis is initiated at the plasma membrane, and proteins are packed into clathrin-coated vesicles before being transported to early and late endosomes. From there, they are either recycled back to the plasma membrane or are degraded in lysosomes. The putative sites where different viruses modify intracellular membranes to assemble their replication complexes are indicated. EAV, equine arteritis virus; FHV, flock house virus; HCV, hepatitis C virus; KUNV, Kunjin virus; MHV, murine hepatitis virus; SARS-CoV, severe acute respiratory syndrome coronavirus; SFV, Semliki Forest virus; TMV, tobacco mosaic virus.
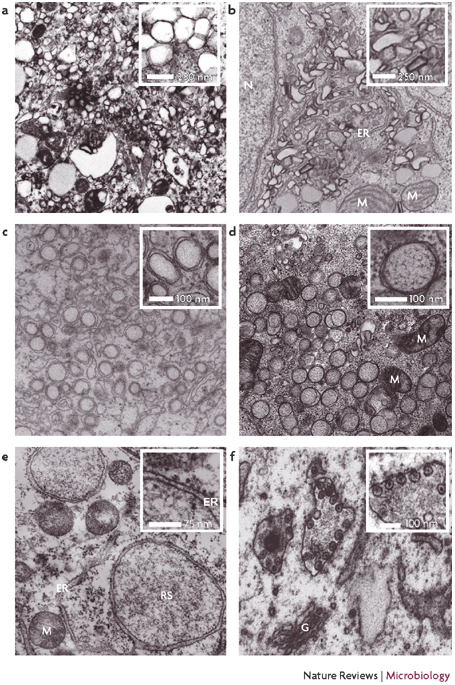
a | Poliovirus-infected HeLa cells fixed 6–8 hours post infection. b | Hepatitis C virus-infected Huh7 cells. c | Dengue virus-infected Huh7 cells fixed 24 hours post infection. d | Severe acute respiratory syndrome coronavirus-infected Vero cells, showing a cluster of double-membrane vesicles (DMVs). The inset shows one DMV at a higher magnification. e | Vaccinia virus-infected cells, showing a replication site that is surrounded by the rough ER. The inset shows the ER membrane at a higher magnification, with the ribosomes on the outer membrane facing the cytoplasm. f | Semliki Forest virus-infected baby hamster kidney cells fixed 3 hours post infection, showing the typical cytopathic vacuoles that are induced upon infection. ER, endoplasmic reticulum; G, Golgi apparatus; M, mitochondrion; N, nucleus; RS, replication site. Part a is courtesy of K. Bienz and D. Egger, University of Basel, Switzerland; parts b and c are courtesy of R. Bartenschlager, University of Heidelberg, Germany; part d is courtesy of E. Snijder, M. Mommas and K. Knoops, Leiden University, The Netherlands; part e is reproduced, with permission, from Ref. © (2005) Blackwell Publishing; and part f is courtesy of T. Ahola and G. Balistrer, University of Helsinki, Finland.
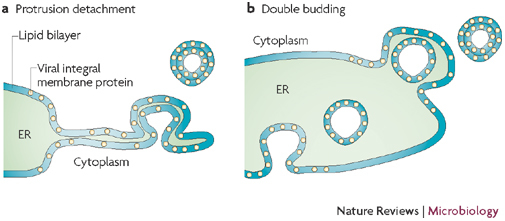
The protrusion and detachment model (a) proposes that part of the endoplasmic reticulum (ER) cisterna starts to bend, pinches off and then seals to form a double-membrane vesicle (DMV). Interactions between the lumenal domains of viral membrane proteins (coloured yellow in the ER membrane) could mediate the tight apposition of the two bilayers and induce curvature. In the double-budding model (b), a single-membrane vesicle buds into the lumen of the ER and then buds out again, and the membrane proteins could mediate inward as well as outward budding. Figure adapted, with permission, from Ref. © (1999) American Society for Microbiology.
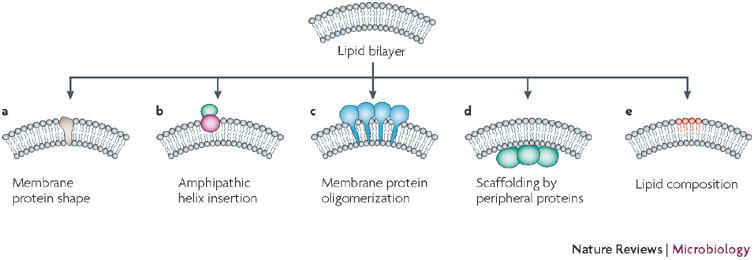
Several mechanisms of membrane deformation by cellular proteins have been described. a | Integration of membrane proteins that have a conical shape induces curvature, as they act like a wedge that is inserted into the membrane. b | Amphipathic helices, stretches of alpha helices that have one polar and one hydrophobic side, are positioned flat on the membrane, with the hydrophobic side dipping into one of the two membrane layers: this causes destabilization of the membrane and membrane bending. c,d | Membrane bending can be induced by oligomerization of proteins that are integrated into, or associated with, cellular membranes. Proteins form a scaffold that makes the membrane bend. e | The lipid composition of a membrane can also induce membrane curvature. In this context, the head group, as well as the acyl chain of the membrane constituents, can have an effect on membrane curvature. Figure adapted, with permission, from Nature Ref. © (2005) Macmillan Publishers Ltd.
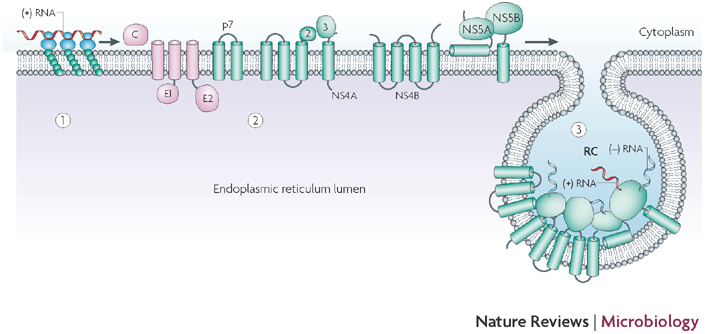
Upon release of viral genomic RNA into the cytoplasm of the infected cell (1), the viral genome is translated into a polyprotein that carries the structural (pink) and non-structural (green) proteins (2). The viral non-structural protein NS4B induces the formation of membrane alterations, which serve as a scaffold for the assembly of the viral replication complex (RC) (3). The RC consists of viral non-structural proteins, viral RNA and host cell factors. Within the induced vesicles, viral RNA is amplified via a negative-strand RNA intermediate. Figure adapted, with permission, from Ref. © (2005) American Society for Microbiology.
Similar articles
-
Alternate, virus-induced membrane rearrangements support positive-strand RNA virus genome replication.Proc Natl Acad Sci U S A. 2004 Aug 3;101(31):11263-8. doi: 10.1073/pnas.0404157101. Epub 2004 Jul 27. Proc Natl Acad Sci U S A. 2004. PMID: 15280537 Free PMC article.
-
Virus factories, double membrane vesicles and viroplasm generated in animal cells.Curr Opin Virol. 2011 Nov;1(5):381-7. doi: 10.1016/j.coviro.2011.09.008. Epub 2011 Oct 12. Curr Opin Virol. 2011. PMID: 22440839 Free PMC article. Review.
-
Ultrastructural Features of Membranous Replication Organelles Induced by Positive-Stranded RNA Viruses.Cells. 2021 Sep 13;10(9):2407. doi: 10.3390/cells10092407. Cells. 2021. PMID: 34572055 Free PMC article. Review.
-
COPII Vesicle Transport Is Required for Rotavirus NSP4 Interaction with the Autophagy Protein LC3 II and Trafficking to Viroplasms.J Virol. 2019 Dec 12;94(1):e01341-19. doi: 10.1128/JVI.01341-19. Print 2019 Dec 12. J Virol. 2019. PMID: 31597778 Free PMC article.
-
Virus-induced double-membrane vesicles.Cell Microbiol. 2015 Jan;17(1):45-50. doi: 10.1111/cmi.12372. Epub 2014 Oct 31. Cell Microbiol. 2015. PMID: 25287059 Free PMC article. Review.
Cited by
-
Ultrastructure of the replication sites of positive-strand RNA viruses.Virology. 2015 May;479-480:418-33. doi: 10.1016/j.virol.2015.02.029. Epub 2015 Mar 6. Virology. 2015. PMID: 25746936 Free PMC article. Review.
-
ACBD3-mediated recruitment of PI4KB to picornavirus RNA replication sites.EMBO J. 2012 Feb 1;31(3):754-66. doi: 10.1038/emboj.2011.429. Epub 2011 Nov 29. EMBO J. 2012. PMID: 22124328 Free PMC article.
-
The triglyceride-synthesizing enzyme diacylglycerol acyltransferase 2 modulates the formation of the hepatitis C virus replication organelle.PLoS Pathog. 2024 Sep 6;20(9):e1012509. doi: 10.1371/journal.ppat.1012509. eCollection 2024 Sep. PLoS Pathog. 2024. PMID: 39241103 Free PMC article.
-
The dependence of viral RNA replication on co-opted host factors.Nat Rev Microbiol. 2011 Dec 19;10(2):137-49. doi: 10.1038/nrmicro2692. Nat Rev Microbiol. 2011. PMID: 22183253 Free PMC article. Review.
-
Usutu virus: an emerging flavivirus in Europe.Viruses. 2015 Jan 19;7(1):219-38. doi: 10.3390/v7010219. Viruses. 2015. PMID: 25606971 Free PMC article. Review.
References
-
- Moss B, et al. Fields Virology. 2001. pp. 2849–2883.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
