

Extracellular ATP reduces HIV-1 transfer from immature dendritic cells to CD4+ T lymphocytes
- PMID: 18373845
- PMCID: PMC2346478
- DOI: 10.1186/1742-4690-5-30
Extracellular ATP reduces HIV-1 transfer from immature dendritic cells to CD4+ T lymphocytes
Abstract
Background: Dendritic cells (DCs) are considered as key mediators of the early events in human immunodeficiency virus type 1 (HIV-1) infection at mucosal sites. Previous studies have shown that surface-bound virions and/or internalized viruses found in endocytic vacuoles of DCs are efficiently transferred to CD4+ T cells. Extracellular adenosine triphosphate (ATP) either secreted or released from necrotic cells induces a distorted maturation of DCs, transiently increases their endocytic capacity and affects their migratory capacity. Knowing that high extracellular ATP concentrations are present in situations of tissue injury and inflammation, we investigated the effect of ATP on HIV-1 transmission from DCs to CD4+ T lymphocytes.
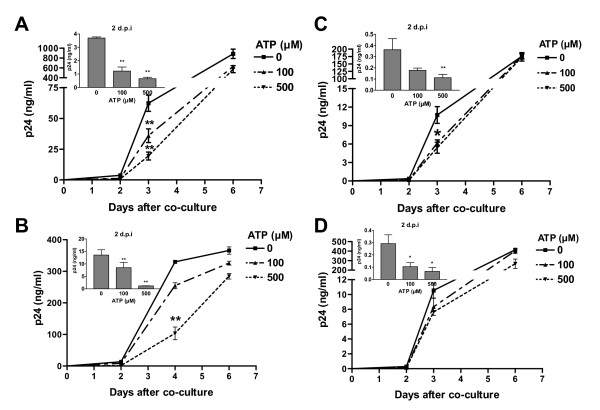
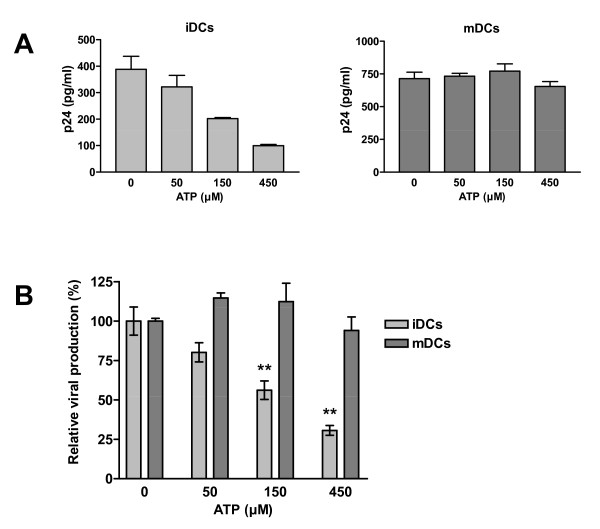
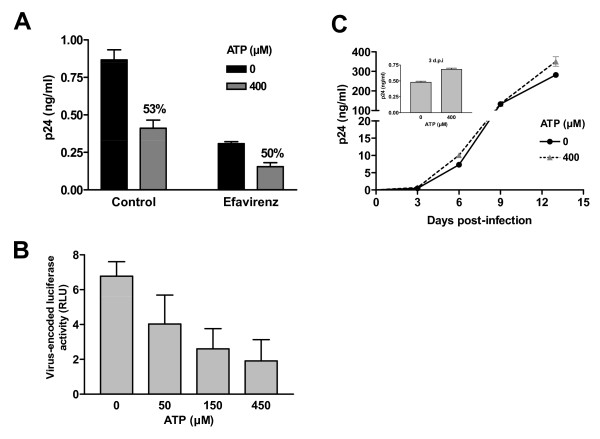
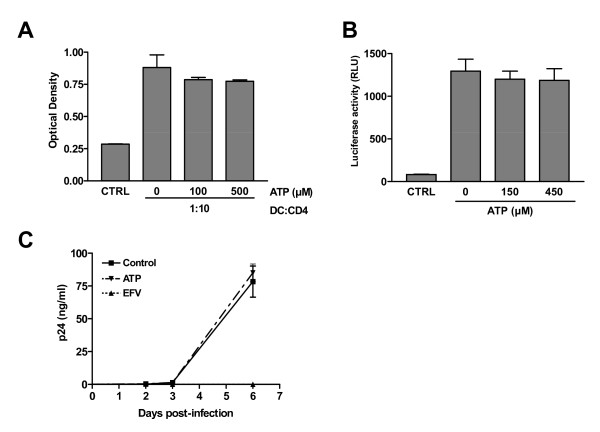
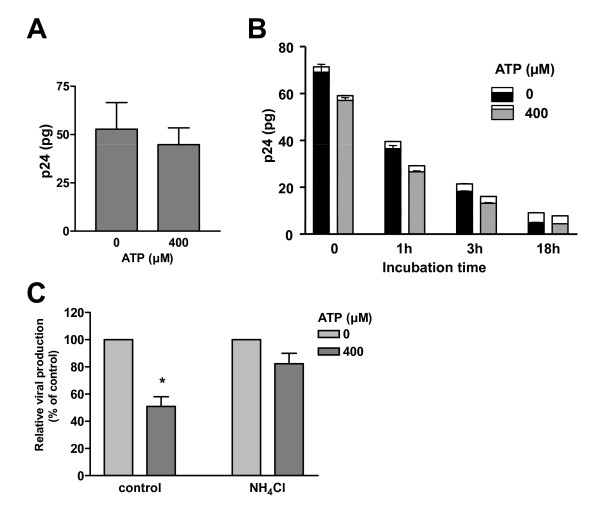
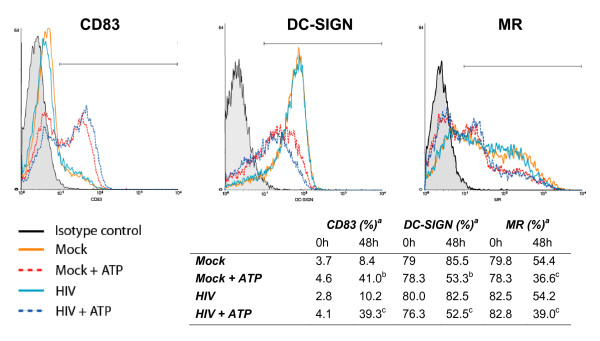
Results: In this study, we show that extracellular ATP reduces HIV-1 transfer from immature monocyte-derived DCs (iDCs) to autologous CD4+ T cells. This observed decrease in viral replication was related to a lower proportion of infected CD4+ T cells following transfer, and was seen with both X4- and R5-tropic isolates of HIV-1. Extracellular ATP had no effect on direct CD4+ T cell infection as well as on productive HIV-1 infection of iDCs. These observations indicate that extracellular ATP affects HIV-1 infection of CD4+ T cells in trans with no effect on de novo virus production by iDCs. Additional experiments suggest that extracellular ATP might modulate the trafficking pathway of internalized virions within iDCs leading to an increased lysosomal degradation, which could be partly responsible for the decreased HIV-1 transmission.
Conclusion: These results suggest that extracellular ATP can act as a factor controlling HIV-1 propagation.
Figures

Similar articles
-
Cholecalciferol modulates the phenotype of differentiated monocyte-derived dendritic cells without altering HIV-1 transfer to CD4+ T cells.Horm Mol Biol Clin Investig. 2019 Sep 20;40(1):/j/hmbci.2019.40.issue-1/hmbci-2019-0003/hmbci-2019-0003.xml. doi: 10.1515/hmbci-2019-0003. Horm Mol Biol Clin Investig. 2019. PMID: 31539353
-
Preferential infection of dendritic cells during human immunodeficiency virus type 1 infection of blood leukocytes.J Virol. 2007 Mar;81(5):2297-306. doi: 10.1128/JVI.01795-06. Epub 2006 Dec 13. J Virol. 2007. PMID: 17166903 Free PMC article.
-
Stimulation of HIV-1 replication in immature dendritic cells in contact with primary CD4 T or B lymphocytes.J Virol. 2010 May;84(9):4172-82. doi: 10.1128/JVI.01567-09. Epub 2010 Feb 10. J Virol. 2010. PMID: 20147388 Free PMC article.
-
Cell-Free versus Cell-to-Cell Infection by Human Immunodeficiency Virus Type 1 and Human T-Lymphotropic Virus Type 1: Exploring the Link among Viral Source, Viral Trafficking, and Viral Replication.J Virol. 2016 Aug 12;90(17):7607-17. doi: 10.1128/JVI.00407-16. Print 2016 Sep 1. J Virol. 2016. PMID: 27334587 Free PMC article. Review.
-
The interaction of immunodeficiency viruses with dendritic cells.Curr Top Microbiol Immunol. 2003;276:1-30. doi: 10.1007/978-3-662-06508-2_1. Curr Top Microbiol Immunol. 2003. PMID: 12797441 Review.
Cited by
-
Purinergic modulation of the immune response to infections.Purinergic Signal. 2022 Mar;18(1):93-113. doi: 10.1007/s11302-021-09838-y. Epub 2022 Jan 8. Purinergic Signal. 2022. PMID: 34997903 Free PMC article. Review.
-
Adenosine and its receptors as therapeutic targets: An overview.Saudi Pharm J. 2013 Jul;21(3):245-53. doi: 10.1016/j.jsps.2012.05.011. Epub 2012 Jun 23. Saudi Pharm J. 2013. PMID: 23960840 Free PMC article.
-
HIV interactions with monocytes and dendritic cells: viral latency and reservoirs.Retrovirology. 2009 Jun 1;6:51. doi: 10.1186/1742-4690-6-51. Retrovirology. 2009. PMID: 19486514 Free PMC article. Review.
-
The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications.Cancers (Basel). 2022 Feb 22;14(5):1116. doi: 10.3390/cancers14051116. Cancers (Basel). 2022. PMID: 35267424 Free PMC article. Review.
-
HIV-1 Trans Infection of CD4(+) T Cells by Professional Antigen Presenting Cells.Scientifica (Cairo). 2013;2013:164203. doi: 10.1155/2013/164203. Epub 2013 May 7. Scientifica (Cairo). 2013. PMID: 24278768 Free PMC article. Review.
References
-
- Smed-Sorensen A, Lore K, Vasudevan J, Louder MK, Andersson J, Mascola JR, Spetz AL, Koup RA. Differential susceptibility to human immunodeficiency virus type 1 infection of myeloid and plasmacytoid dendritic cells. J Virol. 2005;79:8861–8869. doi: 10.1128/JVI.79.14.8861-8869.2005. - DOI - PMC - PubMed
-
- Geijtenbeek TB, Kwon DS, Torensma R, van Vliet SJ, van Duijnhoven GC, Middel J, Cornelissen IL, Nottet HS, KewalRamani VN, Littman DR, Figdor CG, van Kooyk Y. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell. 2000;100:587–597. doi: 10.1016/S0092-8674(00)80694-7. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
