

mGluR1/5-dependent long-term depression requires the regulated ectodomain cleavage of neuronal pentraxin NPR by TACE
- PMID: 18367087
- PMCID: PMC2701195
- DOI: 10.1016/j.neuron.2008.01.010
mGluR1/5-dependent long-term depression requires the regulated ectodomain cleavage of neuronal pentraxin NPR by TACE
Abstract
Matrix metalloproteases (MMPs) play a role in remodeling the extracellular matrix during brain development and have been implicated in synaptic plasticity. Here, we report that a member of the neuronal pentraxin (NP) family, neuronal pentraxin receptor (NPR), undergoes regulated cleavage by the MMP tumor necrosis factor-alpha converting enzyme (TACE). NPR is enriched at excitatory synapses where it associates with AMPA-type glutamate receptors (AMPAR) and enhances synaptogenesis. However, in response to activation of group 1 mGluRs (mGluR1/5), TACE cleaves NPR and releases the pentraxin domain from its N-terminal transmembrane domain. Cleaved NPR rapidly accumulates in endosomes where it colocalizes with AMPAR. This process is necessary for mGluR1/5-dependent LTD in hippocampal and cerebellar synapses. These observations suggest that cleaved NPR functions to "capture" AMPAR for endocytosis and reveal a bifunctional role of NPs in both synapse strengthening and weakening.
Figures

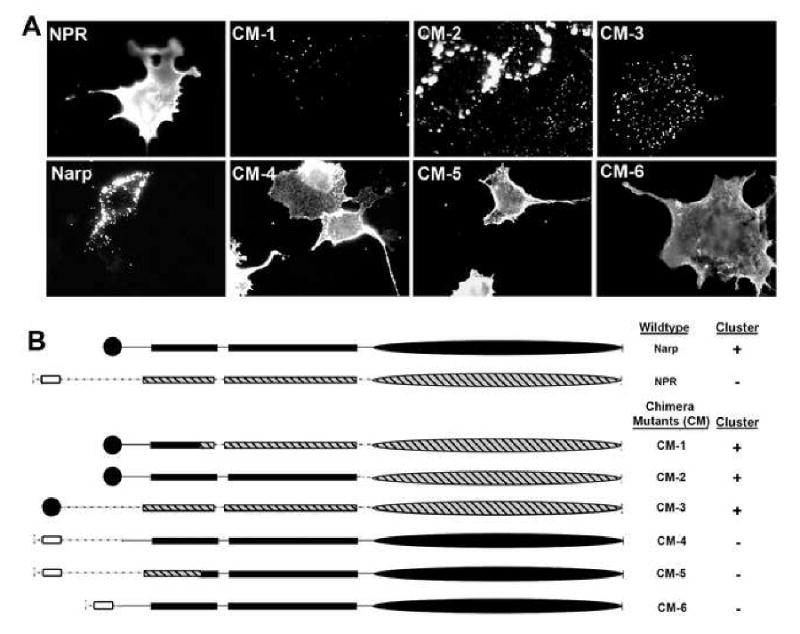

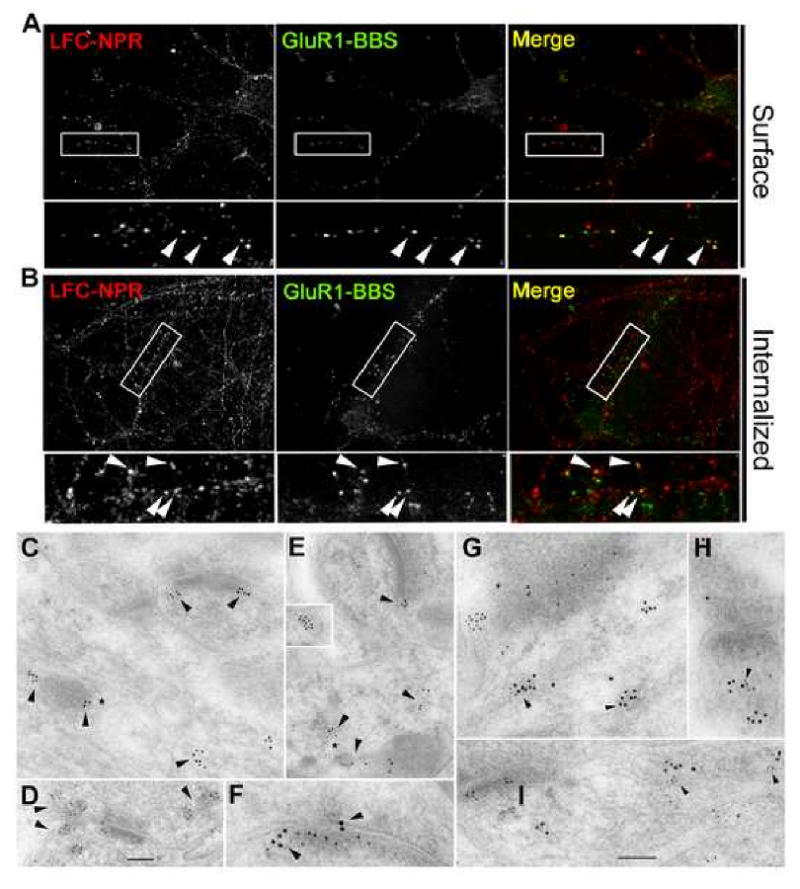
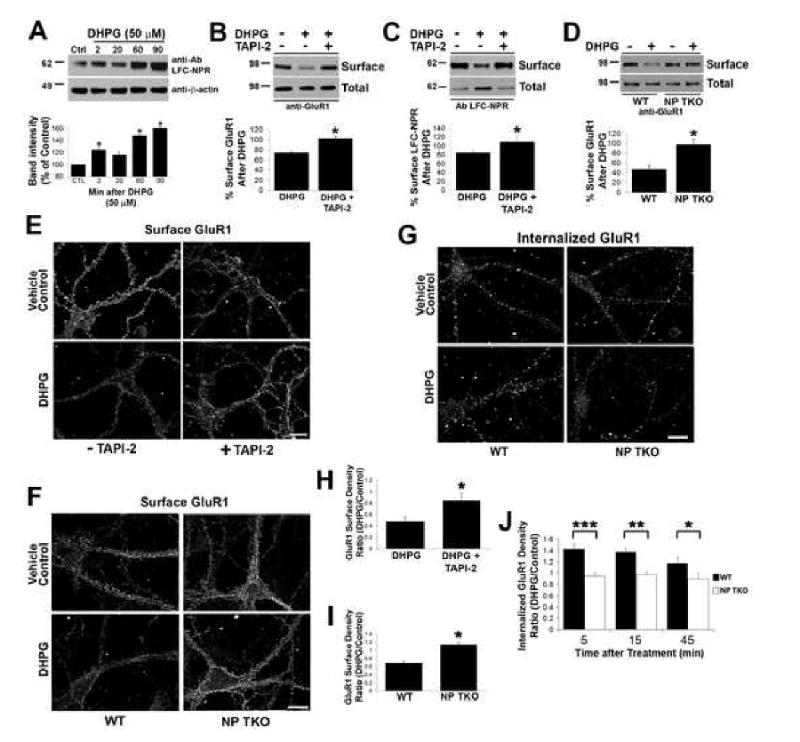

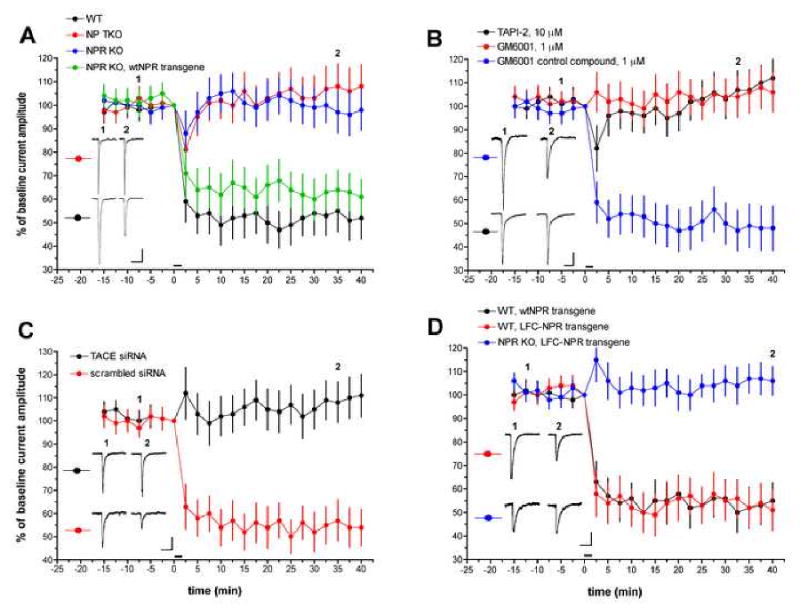

Similar articles
-
Presynaptic Neuronal Pentraxin Receptor Organizes Excitatory and Inhibitory Synapses.J Neurosci. 2017 Feb 1;37(5):1062-1080. doi: 10.1523/JNEUROSCI.2768-16.2016. Epub 2016 Dec 16. J Neurosci. 2017. PMID: 27986928 Free PMC article.
-
Enhancement of long-term depression by soluble amyloid β protein in rat hippocampus is mediated by metabotropic glutamate receptor and involves activation of p38MAPK, STEP and caspase-3.Neuroscience. 2013 Dec 3;253:435-43. doi: 10.1016/j.neuroscience.2013.08.054. Epub 2013 Sep 5. Neuroscience. 2013. PMID: 24012839
-
NMDA receptor activation potentiates inhibitory transmission through GABA receptor-associated protein-dependent exocytosis of GABA(A) receptors.J Neurosci. 2007 Dec 26;27(52):14326-37. doi: 10.1523/JNEUROSCI.4433-07.2007. J Neurosci. 2007. PMID: 18160640 Free PMC article.
-
Do group I metabotropic glutamate receptors mediate LTD?Neurobiol Learn Mem. 2017 Feb;138:85-97. doi: 10.1016/j.nlm.2016.08.010. Epub 2016 Aug 18. Neurobiol Learn Mem. 2017. PMID: 27545442 Review.
-
Roles of phospholipase Cbeta4 in synapse elimination and plasticity in developing and mature cerebellum.Mol Neurobiol. 2001 Feb;23(1):69-82. doi: 10.1385/MN:23:1:69. Mol Neurobiol. 2001. PMID: 11642544 Review.
Cited by
-
Sex-Specific Impact of Fkbp5 on Hippocampal Response to Acute Alcohol Injection: Involvement in Alterations of Metabolism-Related Pathways.Cells. 2023 Dec 31;13(1):89. doi: 10.3390/cells13010089. Cells. 2023. PMID: 38201293 Free PMC article.
-
Matrix metalloproteinase-dependent shedding of intercellular adhesion molecule-5 occurs with long-term potentiation.Neuroscience. 2010 Mar 17;166(2):508-21. doi: 10.1016/j.neuroscience.2009.12.061. Epub 2010 Jan 4. Neuroscience. 2010. PMID: 20045450 Free PMC article.
-
Matrix metalloproteinases, synaptic injury, and multiple sclerosis.Front Psychiatry. 2010 Oct 5;1:130. doi: 10.3389/fpsyt.2010.00130. eCollection 2010. Front Psychiatry. 2010. PMID: 21423441 Free PMC article.
-
Identification of longitudinally dynamic biomarkers in Alzheimer's disease cerebrospinal fluid by targeted proteomics.Mol Neurodegener. 2014 Jun 6;9:22. doi: 10.1186/1750-1326-9-22. Mol Neurodegener. 2014. PMID: 24902845 Free PMC article.
-
MiR-142-3p is a Critical Modulator of TNF-mediated Neuronal Toxicity in Multiple Sclerosis.Curr Neuropharmacol. 2023;21(12):2567-2582. doi: 10.2174/1570159X21666230404103914. Curr Neuropharmacol. 2023. PMID: 37021418 Free PMC article.
References
-
- Aiba A, Kano M, Chen C, Stanton ME, Fox GD, Herrup K, Zwingman TA, Tonegawa S. Deficient cerebellar long-term depression and impaired motor learning in mGluR1 mutant mice. Cell. 1994;79:377–388. - PubMed
-
- Arribas J, Coodly L, Vollmer P, Kishimoto TK, Rose-John S, Massague J. Diverse cell surface protein ectodomains are shed by a system sensitive to metalloprotease inhibitors. J Biol Chem. 1996;271:11376–11382. - PubMed
-
- Bear MF, Linden DJ. The mechanisms and meaning of long-term synaptic depression in the mammalian brain. Baltimore: Johns Hopkins University Press; 2000.
-
- Beckmann G, Hanke J, Bork P, Reich JG. Merging extracellular domains: fold prediction for laminin G-like and amino-terminal thrombospondin-like modules based on homology to pentraxins. J Mol Biol. 1998;275:725–730. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
