

9-phenanthrol inhibits human TRPM4 but not TRPM5 cationic channels
- PMID: 18297105
- PMCID: PMC2438271
- DOI: 10.1038/bjp.2008.38
9-phenanthrol inhibits human TRPM4 but not TRPM5 cationic channels
Abstract
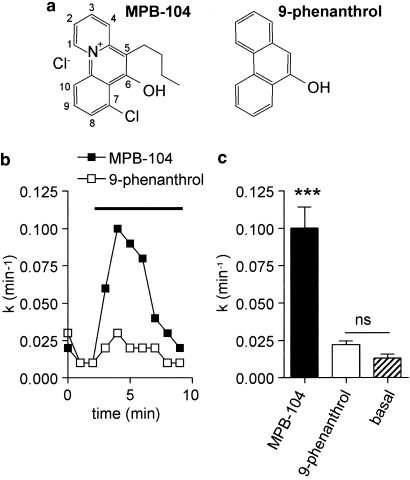
Background and purpose: TRPM4 and TRPM5 are calcium-activated non-selective cation channels with almost identical characteristics. TRPM4 is detected in several tissues including heart, kidney, brainstem, cerebral artery and immune system whereas TRPM5 expression is more restricted. Determination of their roles in physiological processes requires specific pharmacological tools. TRPM4 is inhibited by glibenclamide, a modulator of ATP binding cassette proteins (ABC transporters), such as the cystic fibrosis transmembrane conductance regulator (CFTR). We took advantage of this similarity to investigate the effect of hydroxytricyclic compounds shown to modulate ABC transporters, on TRPM4 and TRPM5.
Experimental approach: Experiments were conducted using HEK-293 cells permanently transfected to express human TRPM4 or TRPM5. Currents were recorded using the whole-cell and inside-out variants of the patch-clamp technique.
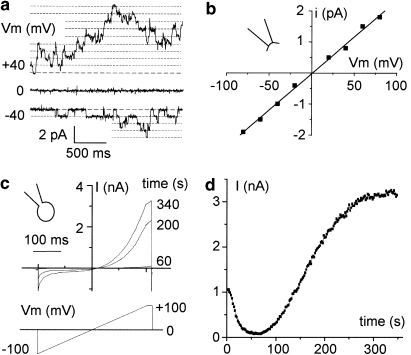
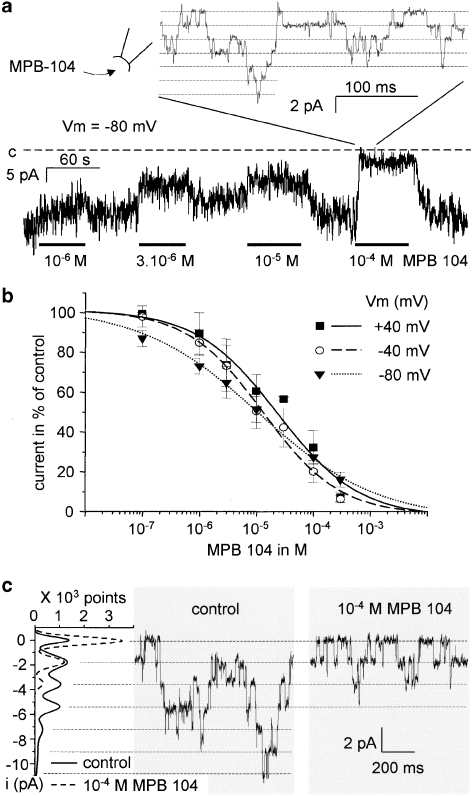
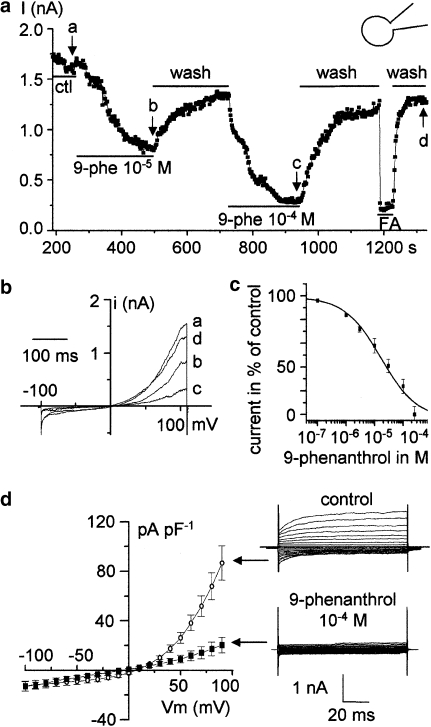
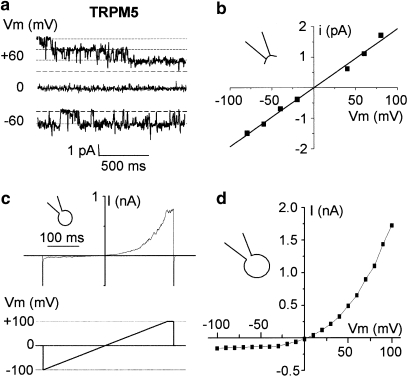
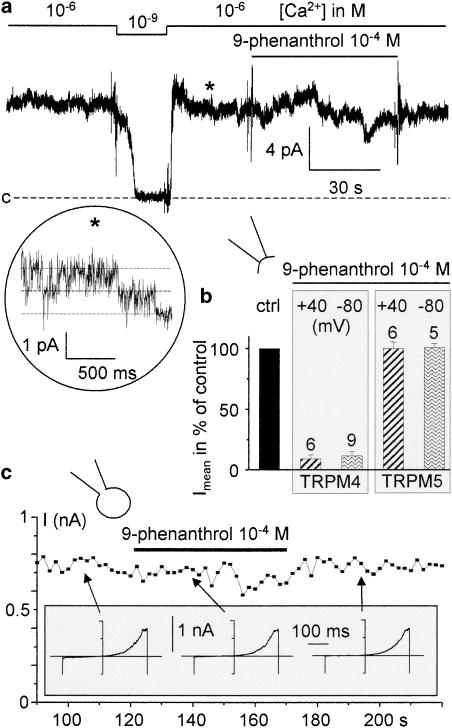
Key results: The CFTR channel activator benzo[c]quinolizinium MPB-104 inhibited TRPM4 current with an IC(50) in the range of 2 x 10(-5) M, with no effect on single-channel conductance. In addition, 9-phenanthrol, lacking the chemical groups necessary for CFTR activation, also reversibly inhibited TRPM4 with a similar IC(50). Channel inhibition was voltage independent. The IC(50) determined in the whole-cell and inside-out experiments were similar, suggesting a direct effect of the molecule. However, 9-phenanthrol was ineffective on TRPM5, the most closely related channel within the TRP protein family.
Conclusions and implications: We identify 9-phenanthrol as a TRPM4 inhibitor, without effects on TRPM5. It could be valuable in investigating the physiological functions of TRPM4, as distinct from those of TRPM5.
Figures

Similar articles
-
Pharmacological inhibition of TRPM4 hyperpolarizes vascular smooth muscle.Am J Physiol Cell Physiol. 2010 Nov;299(5):C1195-202. doi: 10.1152/ajpcell.00269.2010. Epub 2010 Sep 8. Am J Physiol Cell Physiol. 2010. PMID: 20826763 Free PMC article.
-
TRPM4 channel inhibitors 9-phenanthrol and glibenclamide differentially decrease guinea pig detrusor smooth muscle whole-cell cation currents and phasic contractions.Am J Physiol Cell Physiol. 2020 Feb 1;318(2):C406-C421. doi: 10.1152/ajpcell.00055.2019. Epub 2019 Dec 18. Am J Physiol Cell Physiol. 2020. PMID: 31851526 Free PMC article.
-
Development of substituted Benzo[c]quinolizinium compounds as novel activators of the cystic fibrosis chloride channel.J Biol Chem. 1999 Sep 24;274(39):27415-25. doi: 10.1074/jbc.274.39.27415. J Biol Chem. 1999. PMID: 10488073
-
The TRPM4 channel inhibitor 9-phenanthrol.Br J Pharmacol. 2014 Apr;171(7):1600-13. doi: 10.1111/bph.12582. Br J Pharmacol. 2014. PMID: 24433510 Free PMC article. Review.
-
The non-selective monovalent cationic channels TRPM4 and TRPM5.Adv Exp Med Biol. 2011;704:147-71. doi: 10.1007/978-94-007-0265-3_8. Adv Exp Med Biol. 2011. PMID: 21290294 Review.
Cited by
-
9-Phenanthrol and flufenamic acid inhibit calcium oscillations in HL-1 mouse cardiomyocytes.Cell Calcium. 2013 Sep;54(3):193-201. doi: 10.1016/j.ceca.2013.06.003. Epub 2013 Jul 5. Cell Calcium. 2013. PMID: 23831210 Free PMC article.
-
Cellular mechanisms and molecular pathways linking bitter taste receptor signalling to cardiac inflammation, oxidative stress, arrhythmia and contractile dysfunction in heart diseases.Inflammopharmacology. 2023 Feb;31(1):89-117. doi: 10.1007/s10787-022-01086-9. Epub 2022 Dec 6. Inflammopharmacology. 2023. PMID: 36471190 Free PMC article. Review.
-
Role of the TRPM4 Channel in Cardiovascular Physiology and Pathophysiology.Cells. 2018 Jun 15;7(6):62. doi: 10.3390/cells7060062. Cells. 2018. PMID: 29914130 Free PMC article. Review.
-
Thermosensitive TRPV4 channels mediate temperature-dependent microglia movement.Proc Natl Acad Sci U S A. 2021 Apr 27;118(17):e2012894118. doi: 10.1073/pnas.2012894118. Proc Natl Acad Sci U S A. 2021. PMID: 33888579 Free PMC article.
-
A glibenclamide-sensitive TRPM4-mediated component of CA1 excitatory postsynaptic potentials appears in experimental autoimmune encephalomyelitis.Sci Rep. 2022 Apr 9;12(1):6000. doi: 10.1038/s41598-022-09875-6. Sci Rep. 2022. PMID: 35397639 Free PMC article.
References
-
- Becq F. On the discovery and development of CFTR chloride channel activators. Curr Pharm Des. 2006;12:471–484. - PubMed
-
- Becq F, Mettey Y, Gray MA, Galietta LJ, Dormer RL, Merten M, et al. Development of substituted Benzo[c]quinolizinium compounds as novel activators of the cystic fibrosis chloride channel. J Biol Chem. 1999;274:27415–27425. - PubMed
-
- Chen JH, Schulman H, Gardner P. A cAMP-regulated chloride channel in lymphocytes that is affected in cystic fibrosis. Science. 1989;243:657–660. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
