

Motility screen identifies Drosophila IGF-II mRNA-binding protein--zipcode-binding protein acting in oogenesis and synaptogenesis
- PMID: 18282112
- PMCID: PMC2242817
- DOI: 10.1371/journal.pgen.0040036
Motility screen identifies Drosophila IGF-II mRNA-binding protein--zipcode-binding protein acting in oogenesis and synaptogenesis
Abstract
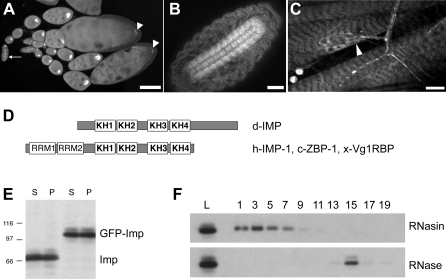
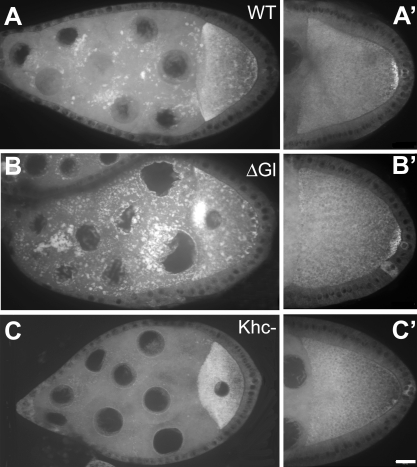
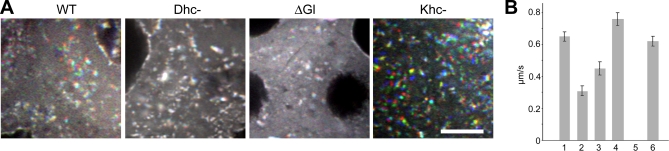
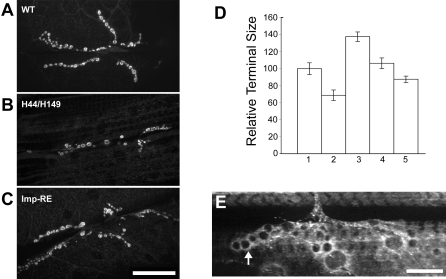
The localization of specific mRNAs can establish local protein gradients that generate and control the development of cellular asymmetries. While all evidence underscores the importance of the cytoskeleton in the transport and localization of RNAs, we have limited knowledge of how these events are regulated. Using a visual screen for motile proteins in a collection of GFP protein trap lines, we identified the Drosophila IGF-II mRNA-binding protein (Imp), an ortholog of Xenopus Vg1 RNA binding protein and chicken zipcode-binding protein. In Drosophila, Imp is part of a large, RNase-sensitive complex that is enriched in two polarized cell types, the developing oocyte and the neuron. Using time-lapse confocal microscopy, we establish that both dynein and kinesin contribute to the transport of GFP-Imp particles, and that regulation of transport in egg chambers appears to differ from that in neurons. In Drosophila, loss-of-function Imp mutations are zygotic lethal, and mutants die late as pharate adults. Imp has a function in Drosophila oogenesis that is not essential, as well as functions that are essential during embryogenesis and later development. Germline clones of Imp mutations do not block maternal mRNA localization or oocyte development, but overexpression of a specific Imp isoform disrupts dorsal/ventral polarity. We report here that loss-of-function Imp mutations, as well as Imp overexpression, can alter synaptic terminal growth. Our data show that Imp is transported to the neuromuscular junction, where it may modulate the translation of mRNA targets. In oocytes, where Imp function is not essential, we implicate a specific Imp domain in the establishment of dorsoventral polarity.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
Similar articles
-
In vivo analyses of cytoplasmic transport and cytoskeletal organization during Drosophila oogenesis: characterization of a multi-step anterior localization pathway.Development. 1998 Sep;125(18):3655-66. doi: 10.1242/dev.125.18.3655. Development. 1998. PMID: 9716531
-
Identification of genes that influence gurken expression.Fly (Austin). 2007 Sep-Oct;1(5):259-67. doi: 10.4161/fly.5246. Epub 2007 Sep 4. Fly (Austin). 2007. PMID: 18836312
-
U7 snRNA mutations in Drosophila block histone pre-mRNA processing and disrupt oogenesis.RNA. 2006 Mar;12(3):396-409. doi: 10.1261/rna.2270406. RNA. 2006. PMID: 16495235 Free PMC article.
-
Localization, anchoring and translational control of oskar, gurken, bicoid and nanos mRNA during Drosophila oogenesis.Fly (Austin). 2009 Jan-Mar;3(1):15-28. doi: 10.4161/fly.3.1.7751. Epub 2009 Jan 2. Fly (Austin). 2009. PMID: 19182536 Review.
-
Signaling between somatic follicle cells and the germline patterns the egg and embryo of Drosophila.Curr Top Dev Biol. 2020;140:55-86. doi: 10.1016/bs.ctdb.2019.10.004. Epub 2019 Nov 19. Curr Top Dev Biol. 2020. PMID: 32591083 Review.
Cited by
-
Insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) in hematological diseases.Mol Med. 2024 Sep 28;30(1):165. doi: 10.1186/s10020-024-00936-2. Mol Med. 2024. PMID: 39342091 Free PMC article. Review.
-
Identification of Genes Involved in the Differentiation of R7y and R7p Photoreceptor Cells in Drosophila.G3 (Bethesda). 2020 Nov 5;10(11):3949-3958. doi: 10.1534/g3.120.401370. G3 (Bethesda). 2020. PMID: 32972998 Free PMC article.
-
S6 kinase localizes to the presynaptic active zone and functions with PDK1 to control synapse development.J Cell Biol. 2011 Sep 19;194(6):921-35. doi: 10.1083/jcb.201101042. J Cell Biol. 2011. PMID: 21930778 Free PMC article.
-
Neuronal polarity in Drosophila: sorting out axons and dendrites.Dev Neurobiol. 2011 Jun;71(6):419-29. doi: 10.1002/dneu.20836. Dev Neurobiol. 2011. PMID: 21557498 Free PMC article. Review.
-
Insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs): post-transcriptional drivers of cancer progression?Cell Mol Life Sci. 2013 Aug;70(15):2657-75. doi: 10.1007/s00018-012-1186-z. Epub 2012 Oct 16. Cell Mol Life Sci. 2013. PMID: 23069990 Free PMC article. Review.
References
-
- Lipshitz HD, Smibert CA. Mechanisms of RNA localization and translational regulation. Curr Opin Genet Dev. 2000;10:476–488. - PubMed
-
- Czaplinski K, Singer RH. Pathways for mRNA localization in the cytoplasm. Trends Biochem Sci. 2006;31:687–693. - PubMed
-
- Litman P, Barg J, Rindzoonski L, Ginzburg I. Subcellular localization of tau mRNA in differentiating neuronal cell culture: implications for neuronal polarity. Neuron. 1993;10:627–638. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
