

Bunched, the Drosophila homolog of the mammalian tumor suppressor TSC-22, promotes cellular growth
- PMID: 18226226
- PMCID: PMC2253523
- DOI: 10.1186/1471-213X-8-10
Bunched, the Drosophila homolog of the mammalian tumor suppressor TSC-22, promotes cellular growth
Abstract
Background: Transforming Growth Factor-beta1 stimulated clone-22 (TSC-22) is assumed to act as a negative growth regulator and tumor suppressor. TSC-22 belongs to a family of putative transcription factors encoded by four distinct loci in mammals. Possible redundancy among the members of the TSC-22/Dip/Bun protein family complicates a genetic analysis. In Drosophila, all proteins homologous to the TSC-22/Dip/Bun family members are derived from a single locus called bunched (bun).
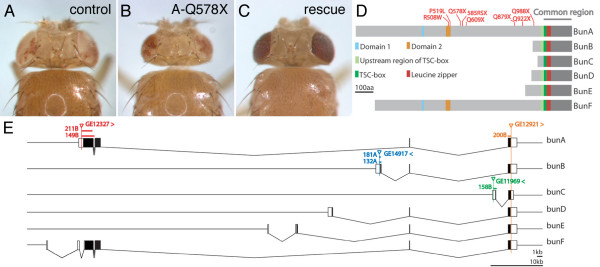
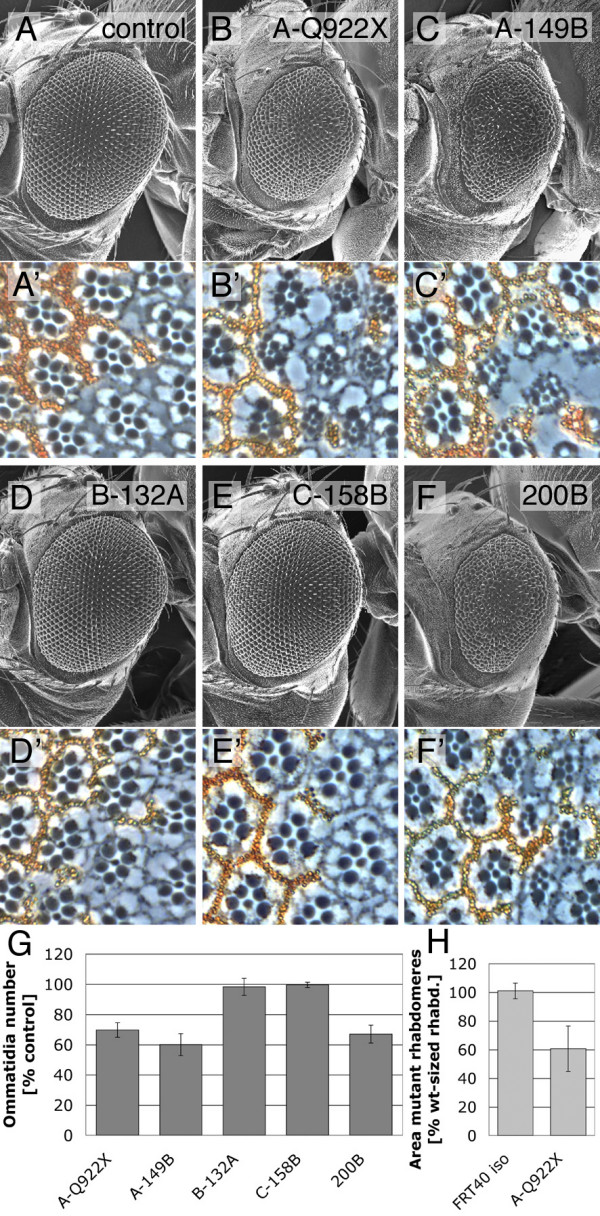
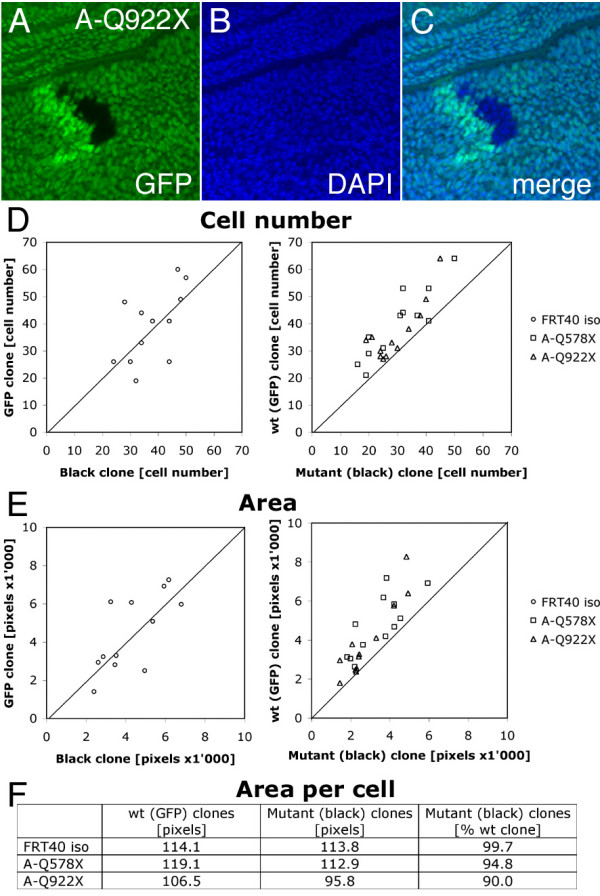
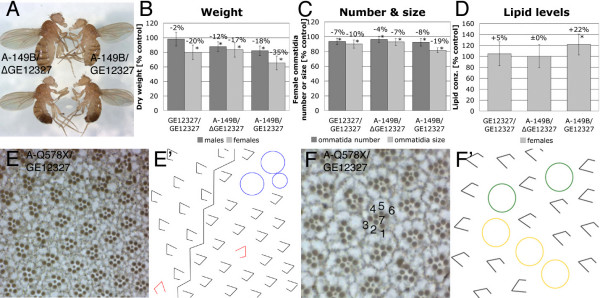
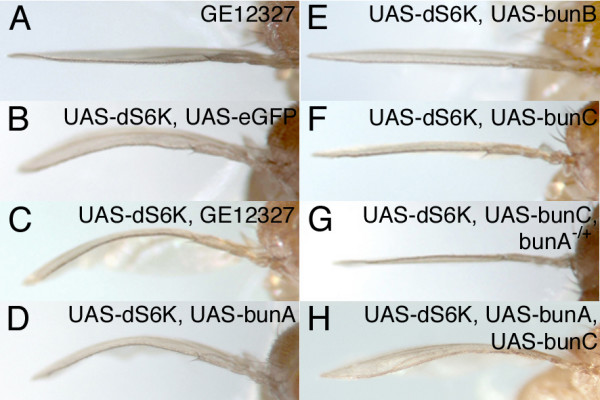
Results: We have identified bun in an unbiased genetic screen for growth regulators in Drosophila. Rather unexpectedly, bun mutations result in a growth deficit. Under standard conditions, only the long protein isoform BunA - but not the short isoforms BunB and BunC - is essential and affects growth. Whereas reducing bunA function diminishes cell number and cell size, overexpression of the short isoforms BunB and BunC antagonizes bunA function.
Conclusion: Our findings establish a growth-promoting function of Drosophila BunA. Since the published studies on mammalian systems have largely neglected the long TSC-22 protein version, we hypothesize that the long TSC-22 protein is a functional homolog of BunA in growth regulation, and that it is antagonized by the short TSC-22 protein.
Figures
Similar articles
-
Madm (Mlf1 adapter molecule) cooperates with Bunched A to promote growth in Drosophila.J Biol. 2010;9(1):9. doi: 10.1186/jbiol216. Epub 2010 Feb 11. J Biol. 2010. PMID: 20149264 Free PMC article.
-
The Drosophila homolog of human tumor suppressor TSC-22 promotes cellular growth, proliferation, and survival.Proc Natl Acad Sci U S A. 2008 Apr 8;105(14):5414-9. doi: 10.1073/pnas.0800945105. Epub 2008 Mar 28. Proc Natl Acad Sci U S A. 2008. PMID: 18375761 Free PMC article.
-
The Drosophila bunched gene is a homologue of the growth factor stimulated mammalian TSC-22 sequence and is required during oogenesis.Mech Dev. 1997 Jul;65(1-2):197-208. doi: 10.1016/s0925-4773(97)00080-4. Mech Dev. 1997. PMID: 9256356
-
Tuberous sclerosis gene products in proliferation control.Mutat Res. 2001 Jul;488(3):233-9. doi: 10.1016/s1383-5742(01)00058-8. Mutat Res. 2001. PMID: 11397651 Review.
-
The tuberous sclerosis complex (TSC) pathway and mechanism of size control.Biochem Soc Trans. 2003 Jun;31(Pt 3):584-6. doi: 10.1042/bst0310584. Biochem Soc Trans. 2003. PMID: 12773160 Review.
Cited by
-
Comparative Genomics Sheds Light on the Convergent Evolution of Miniaturized Wasps.Mol Biol Evol. 2021 Dec 9;38(12):5539-5554. doi: 10.1093/molbev/msab273. Mol Biol Evol. 2021. PMID: 34515790 Free PMC article.
-
Integrated genomics approach to identify biologically relevant alterations in fewer samples.BMC Genomics. 2015 Nov 14;16:936. doi: 10.1186/s12864-015-2138-4. BMC Genomics. 2015. PMID: 26572163 Free PMC article.
-
Identification and characterization of genes required for compensatory growth in Drosophila.Genetics. 2011 Dec;189(4):1309-26. doi: 10.1534/genetics.111.132993. Epub 2011 Sep 16. Genetics. 2011. PMID: 21926302 Free PMC article.
-
TSC-22 contributes to hematopoietic precursor cell proliferation and repopulation and is epigenetically silenced in large granular lymphocyte leukemia.Blood. 2009 May 28;113(22):5558-67. doi: 10.1182/blood-2009-02-205732. Epub 2009 Mar 27. Blood. 2009. PMID: 19329776 Free PMC article.
-
The RNA-binding proteins FMR1, rasputin and caprin act together with the UBA protein lingerer to restrict tissue growth in Drosophila melanogaster.PLoS Genet. 2013;9(7):e1003598. doi: 10.1371/journal.pgen.1003598. Epub 2013 Jul 11. PLoS Genet. 2013. PMID: 23874212 Free PMC article.
References
-
- Shibanuma M, Kuroki T, Nose K. Isolation of a gene encoding a putative leucine zipper structure that is induced by transforming growth factor beta 1 and other growth factors. J Biol Chem. 1992;267:10219–10224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
