

High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds
- PMID: 18162586
- PMCID: PMC2259091
- DOI: 10.1104/pp.107.113738
High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds
Abstract
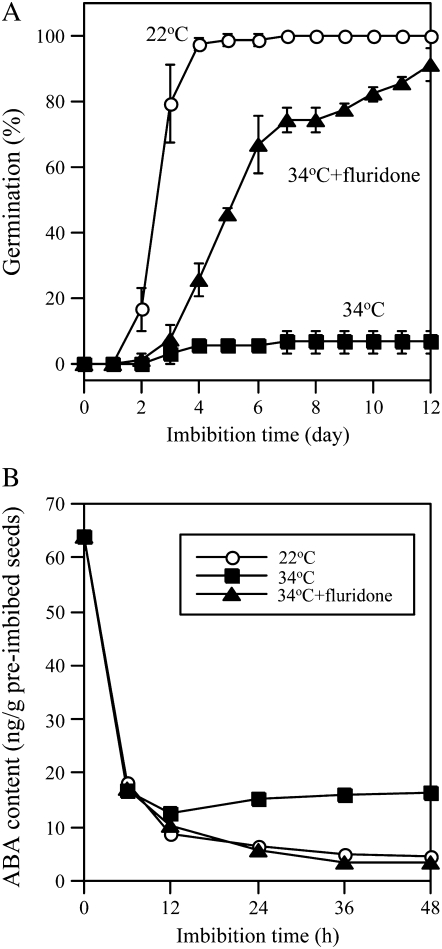
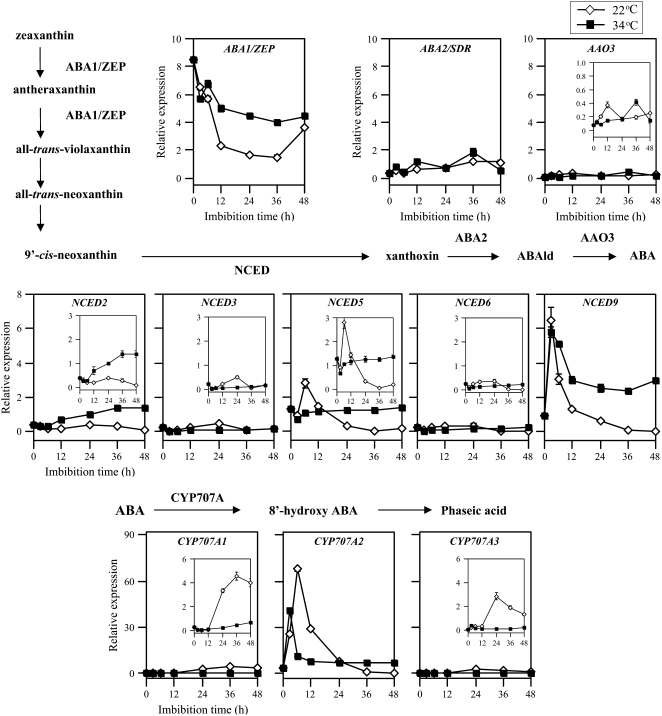
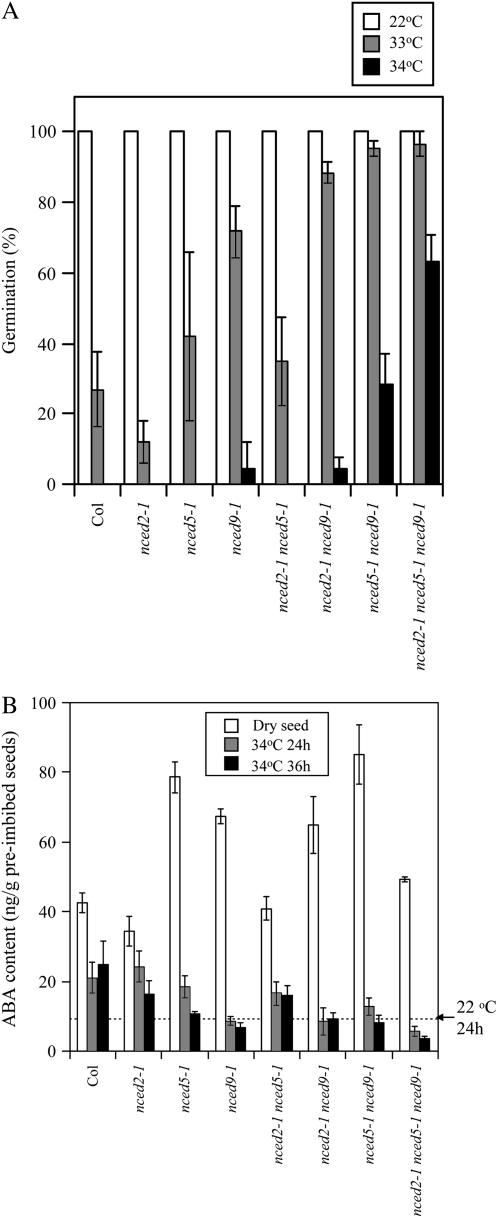
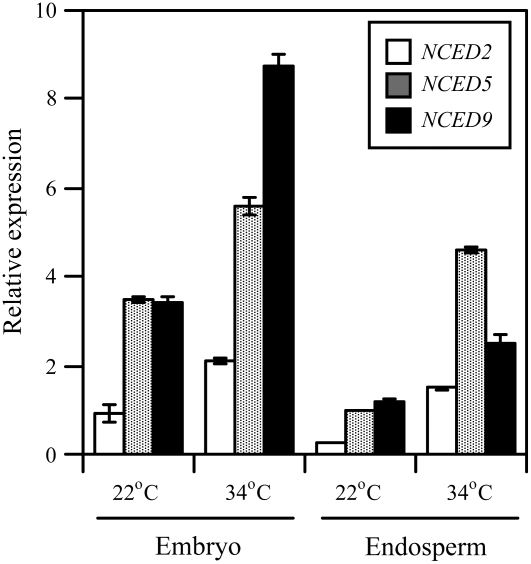
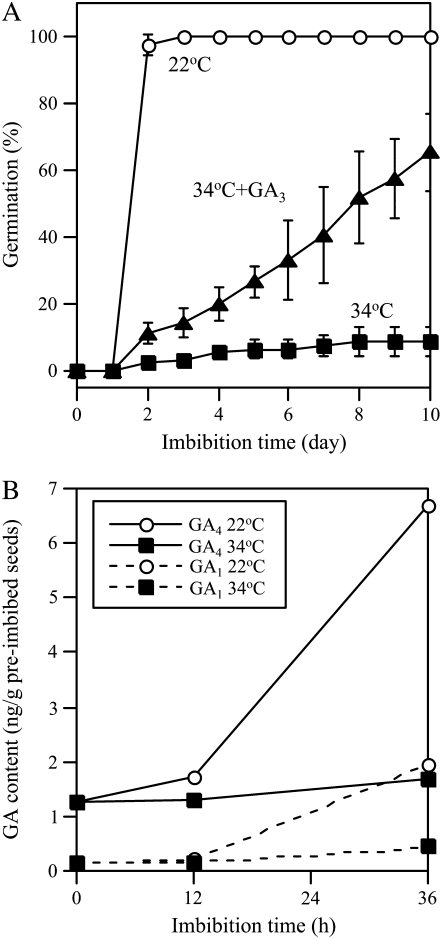
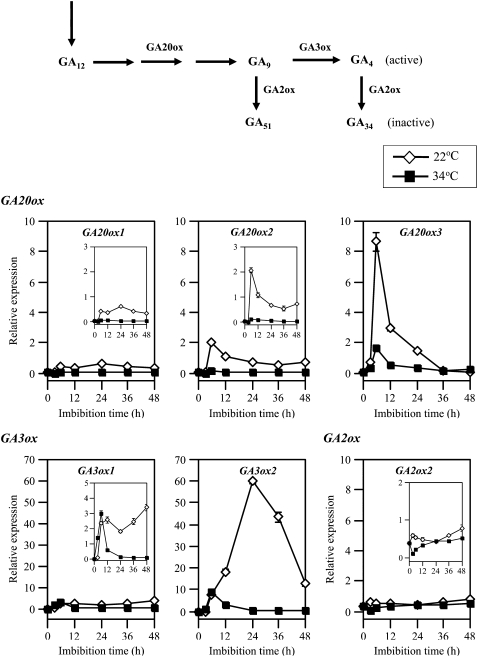
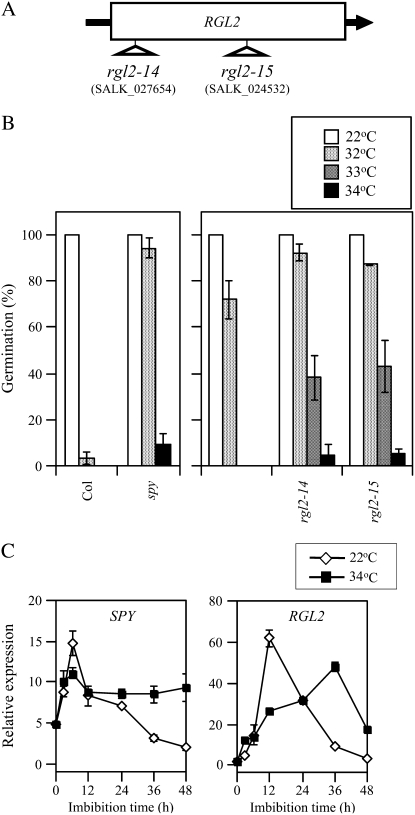
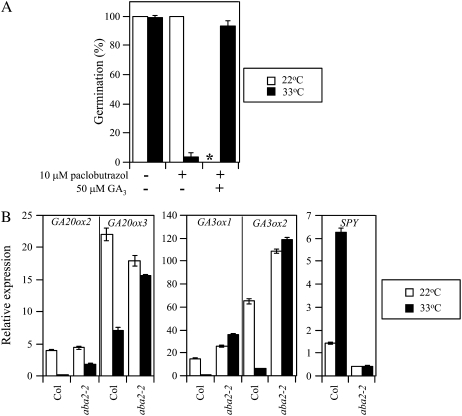
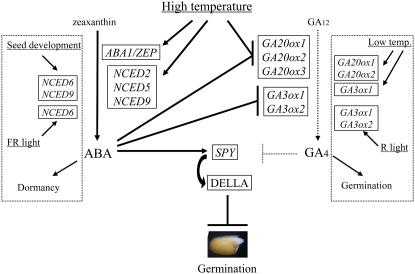
Suppression of seed germination at supraoptimal high temperature (thermoinhibiton) during summer is crucial for Arabidopsis (Arabidopsis thaliana) to establish vegetative and reproductive growth in appropriate seasons. Abscisic acid (ABA) and gibberellins (GAs) are well known to be involved in germination control, but it remains unknown how these hormone actions (metabolism and responsiveness) are altered at high temperature. Here, we show that ABA levels in imbibed seeds are elevated at high temperature and that this increase is correlated with up-regulation of the zeaxanthin epoxidase gene ABA1/ZEP and three 9-cis-epoxycarotenoid dioxygenase genes, NCED2, NCED5, and NCED9. Reverse-genetic studies show that NCED9 plays a major and NCED5 and NCED2 play relatively minor roles in high temperature-induced ABA synthesis and germination inhibition. We also show that bioactive GAs stay at low levels at high temperature, presumably through suppression of GA 20-oxidase genes, GA20ox1, GA20ox2, and GA20ox3, and GA 3-oxidase genes, GA3ox1 and GA3ox2. Thermoinhibition-tolerant germination of loss-of-function mutants of GA negative regulators, SPINDLY (SPY) and RGL2, suggests that repression of GA signaling is required for thermoinibition. Interestingly, ABA-deficient aba2-2 mutant seeds show significant expression of GA synthesis genes and repression of SPY expression even at high temperature. In addition, the thermoinhibition-resistant germination phenotype of aba2-1 seeds is suppressed by a GA biosynthesis inhibitor, paclobutrazol. We conclude that high temperature stimulates ABA synthesis and represses GA synthesis and signaling through the action of ABA in Arabidopsis seeds.
Figures
Similar articles
-
AtPER1 enhances primary seed dormancy and reduces seed germination by suppressing the ABA catabolism and GA biosynthesis in Arabidopsis seeds.Plant J. 2020 Jan;101(2):310-323. doi: 10.1111/tpj.14542. Epub 2019 Oct 22. Plant J. 2020. PMID: 31536657
-
The Arabidopsis MYB96 transcription factor plays a role in seed dormancy.Plant Mol Biol. 2015 Mar;87(4-5):371-81. doi: 10.1007/s11103-015-0283-4. Epub 2015 Jan 24. Plant Mol Biol. 2015. PMID: 25616734
-
Regulation of hormone metabolism in Arabidopsis seeds: phytochrome regulation of abscisic acid metabolism and abscisic acid regulation of gibberellin metabolism.Plant J. 2006 Nov;48(3):354-66. doi: 10.1111/j.1365-313X.2006.02881.x. Epub 2006 Sep 29. Plant J. 2006. PMID: 17010113
-
ABSCISIC ACID INSENSITIVE 5 mediates light-ABA/gibberellin crosstalk networks during seed germination.J Exp Bot. 2022 Aug 11;73(14):4674-4682. doi: 10.1093/jxb/erac200. J Exp Bot. 2022. PMID: 35522989 Review.
-
Interaction of light and hormone signals in germinating seeds.Plant Mol Biol. 2009 Mar;69(4):463-72. doi: 10.1007/s11103-008-9429-y. Epub 2008 Nov 25. Plant Mol Biol. 2009. PMID: 19031046 Review.
Cited by
-
The zinc finger transcription factor SlZFP2 negatively regulates abscisic acid biosynthesis and fruit ripening in tomato.Plant Physiol. 2015 Mar;167(3):931-49. doi: 10.1104/pp.114.255174. Epub 2015 Jan 30. Plant Physiol. 2015. PMID: 25637453 Free PMC article.
-
Enhancement in Seed Priming-Induced Starch Degradation of Rice Seed Under Chilling Stress via GA-Mediated α-Amylase Expression.Rice (N Y). 2022 Mar 27;15(1):19. doi: 10.1186/s12284-022-00567-3. Rice (N Y). 2022. PMID: 35344097 Free PMC article.
-
Seed germination and vigor: ensuring crop sustainability in a changing climate.Heredity (Edinb). 2022 Jun;128(6):450-459. doi: 10.1038/s41437-022-00497-2. Epub 2022 Jan 10. Heredity (Edinb). 2022. PMID: 35013549 Free PMC article. Review.
-
Control of abscisic acid catabolism and abscisic acid homeostasis is important for reproductive stage stress tolerance in cereals.Plant Physiol. 2011 Jun;156(2):647-62. doi: 10.1104/pp.111.176164. Epub 2011 Apr 18. Plant Physiol. 2011. PMID: 21502188 Free PMC article.
-
Distinct gibberellin functions during and after grapevine bud dormancy release.J Exp Bot. 2018 Mar 24;69(7):1635-1648. doi: 10.1093/jxb/ery022. J Exp Bot. 2018. PMID: 29385616 Free PMC article.
References
-
- Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, Van Der Straeten D, Peng J, Harberd NP (2006) Integration of plant responses to environmentally activated phytohormonal signals. Science 311 91–94 - PubMed
-
- Ali-Rachedi S, Bouinot D, Wagner MH, Bonnet M, Sotta B, Grappin P, Jullien M (2004) Changes in endogenous abscisic acid levels during dormancy release and maintenance of mature seeds: studies with the Cape Verde Islands ecotype, the dormant model of Arabidopsis thaliana. Planta 219 479–488 - PubMed
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
