

Arabidopsis FIERY1, XRN2, and XRN3 are endogenous RNA silencing suppressors
- PMID: 17993620
- PMCID: PMC2174888
- DOI: 10.1105/tpc.107.055319
Arabidopsis FIERY1, XRN2, and XRN3 are endogenous RNA silencing suppressors
Abstract
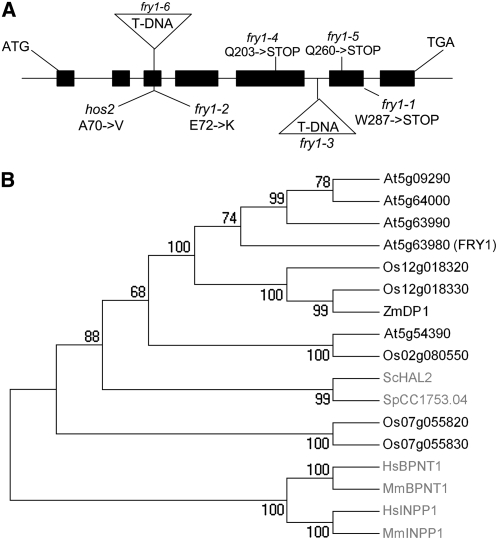
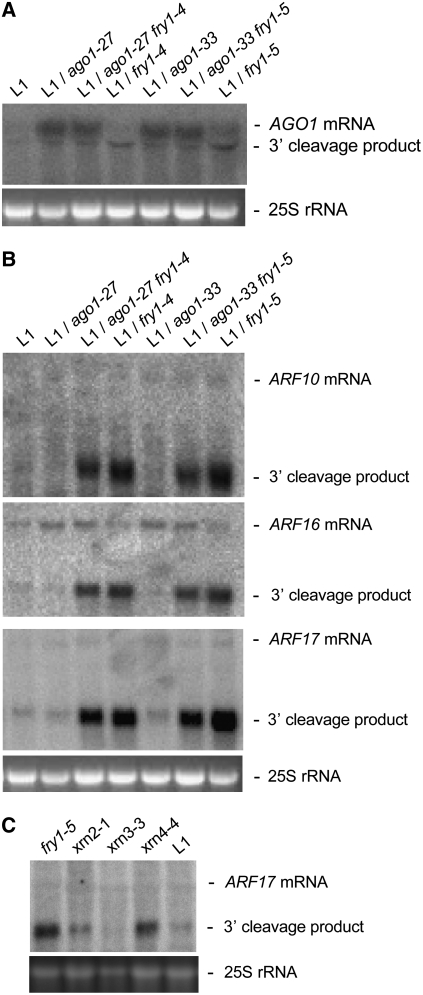
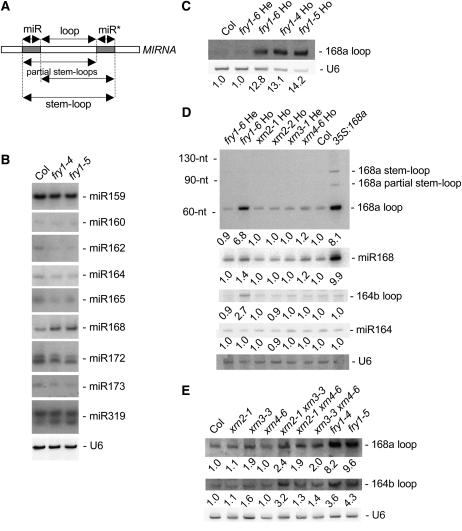
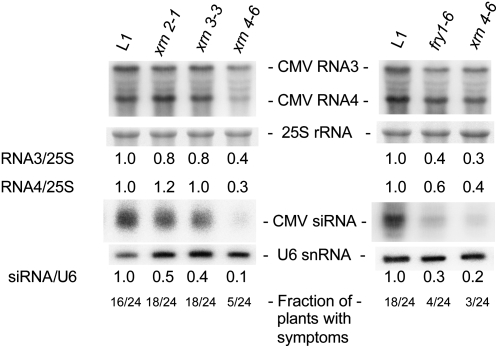
The eukaryotic defense response posttranscriptional gene silencing (PTGS) is directed by short-interfering RNAs and thwarts invading nucleic acids via the RNA slicing activity of conserved ARGONAUTE (AGO) proteins. PTGS can be counteracted by exogenous or endogenous suppressors, including the cytoplasmic exoribonuclease XRN4, which also degrades microRNA (miRNA)-guided mRNA cleavage products but does not play an obvious role in development. Here, we show that the nuclear exoribonucleases XRN2 and XRN3 are endogenous PTGS suppressors. We also identify excised MIRNA loops as templates for XRN2 and XRN3 and show that XRN3 is critical for proper development. Independently, we identified the nucleotidase/phosphatase FIERY1 (FRY1) as an endogenous PTGS suppressor through a suppressor screen in a hypomorphic ago1 genetic background. FRY1 is one of six Arabidopsis thaliana orthologs of yeast Hal2. Yeast hal2 mutants overaccumulate 3'-phosphoadenosine 5'-phosphate, which suppresses the 5'-->3' exoribonucleases Xrn1 and Rat1. fry1 mutant plants recapitulate developmental and molecular characteristics of xrn mutants and likely restore PTGS in ago1 hypomorphic mutants by corepressing XRN2, XRN3, and XRN4, thus increasing RNA silencing triggers. We anticipate that screens incorporating partially compromised silencing components will uncover additional PTGS suppressors that may not be revealed using robust silencing systems.
Figures




Similar articles
-
FIERY1 promotes microRNA accumulation by suppressing rRNA-derived small interfering RNAs in Arabidopsis.Nat Commun. 2019 Sep 27;10(1):4424. doi: 10.1038/s41467-019-12379-z. Nat Commun. 2019. PMID: 31562313 Free PMC article.
-
The bifunctional abiotic stress signalling regulator and endogenous RNA silencing suppressor FIERY1 is required for lateral root formation.Plant Cell Environ. 2010 Dec;33(12):2180-90. doi: 10.1111/j.1365-3040.2010.02218.x. Epub 2010 Sep 28. Plant Cell Environ. 2010. PMID: 20807376
-
Activity and roles of Arabidopsis thaliana XRN family exoribonucleases in noncoding RNA pathways.J Plant Res. 2017 Jan;130(1):25-31. doi: 10.1007/s10265-016-0887-z. Epub 2016 Dec 17. J Plant Res. 2017. PMID: 27988817 Review.
-
Second-Site Mutagenesis of a Hypomorphic argonaute1 Allele Identifies SUPERKILLER3 as an Endogenous Suppressor of Transgene Posttranscriptional Gene Silencing.Plant Physiol. 2015 Oct;169(2):1266-74. doi: 10.1104/pp.15.00585. Epub 2015 Aug 18. Plant Physiol. 2015. PMID: 26286717 Free PMC article.
-
XRN 5'→3' exoribonucleases: structure, mechanisms and functions.Biochim Biophys Acta. 2013 Jun-Jul;1829(6-7):590-603. doi: 10.1016/j.bbagrm.2013.03.005. Epub 2013 Mar 19. Biochim Biophys Acta. 2013. PMID: 23517755 Free PMC article. Review.
Cited by
-
Recent advances in the regulation of plant miRNA biogenesis.RNA Biol. 2021 Dec;18(12):2087-2096. doi: 10.1080/15476286.2021.1899491. Epub 2021 Mar 17. RNA Biol. 2021. PMID: 33666136 Free PMC article. Review.
-
RNA decay is an antiviral defense in plants that is counteracted by viral RNA silencing suppressors.PLoS Pathog. 2018 Aug 3;14(8):e1007228. doi: 10.1371/journal.ppat.1007228. eCollection 2018 Aug. PLoS Pathog. 2018. PMID: 30075014 Free PMC article.
-
Development of strategies for genetic manipulation and fine-tuning of a chloroplast retrograde signal 3'-phosphoadenosine 5'-phosphate.Plant Direct. 2018 Jan 9;2(1):e00031. doi: 10.1002/pld3.31. eCollection 2018 Jan. Plant Direct. 2018. PMID: 31245680 Free PMC article.
-
RNAi-dependent and independent control of LINE1 accumulation and mobility in mouse embryonic stem cells.PLoS Genet. 2013 Nov;9(11):e1003791. doi: 10.1371/journal.pgen.1003791. Epub 2013 Nov 7. PLoS Genet. 2013. Retraction in: PLoS Genet. 2015 Sep 03;11(9):e1005519. doi: 10.1371/journal.pgen.1005519 PMID: 24244175 Free PMC article. Retracted.
-
Modulation of sulfur metabolism enables efficient glucosinolate engineering.BMC Biotechnol. 2011 Jan 31;11:12. doi: 10.1186/1472-6750-11-12. BMC Biotechnol. 2011. PMID: 21281472 Free PMC article.
References
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657. - PubMed
-
- Ambros, V., and Chen, X. (2007). The regulation of genes and genomes by small RNAs. Development 134 1635–1641. - PubMed
-
- Anandalakshmi, R., Marathe, R., Ge, X., Herr, J.M., Jr., Mau, C., Mallory, A., Pruss, G., Bowman, L., and Vance, V.B. (2000). A calmodulin-related protein that suppresses posttranscriptional gene silencing in plants. Science 290 142–144. - PubMed
-
- Bartel, D.P. (2004). MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 116 281–297. - PubMed
-
- Baulcombe, D. (2004). RNA silencing in plants. Nature 431 356–363. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
